For more than 130 years dinosaurs have been classified into two distinct orders, the saurischians and the ornithischians. But as it is common in biological sciences, every theory is true until the opposite is proved. A new study has called into question classical dinosaur classification, destroying and redistributing some of the different dinosaur groups. Even if this new hypothesis isn’t 100% sure yet, in this entry we’ll explain what this dinosaur reordering consists in.
TRADITIONAL DINOSAUR CLASSIFICATION
Since the XIX century, dinosaurs have been divided into two large orders based on their hip anatomy. The order Saurischia (lizard-hipped) includes theropods(carnivorous dinosaurs and current birds) and sauropodomorphs(large, long-necked herbivores); the order Ornithischia (bird-hipped) includes ornithopods(herbivorous and duck-billed dinosaurs), marginocephalians(dinosaurs with horns and hardened skulls) and thyreophorans(armored dinosaurs).
Traditional dinosaur evolutionary tree by Zureks, with the two different hip morphologies at the bottom.
Yet, this classification doesn’t have the last word. Palaeontology is an extremely volatile science, as with each new discovery, you can dismantle everything you knew at that moment, even if it’s a centenary-old hypothesis. This is what has recently happened with dinosaurs.
THE RISE OF A NEW HYPOTHESIS
A new study published in March 2017, has caused the reconsideration of traditional dinosaur classification. Many previous studies assumed the Saurichia/Ornithischia classification as true and so, the used characters and taxons were all focussed on this classification. However, this new study has pioneered in many aspects:
It includes a larger number of species and taxons (many more than in previous investigations).
Previous studies gave more importance to basal theropod and sauropodomorph dinosaurs (traditional saurischians), as they were the first dinosaurs to diversify, including few basal ornithischians.
It has also included many dinosauromorph archosaurs (non-dinosaur taxons).
Older studies had assumed many ornithischian characters to be symplesiomorphies (ancestral characters of all dinosaurs) and they only focused on a few synapomorphies (characters found in a monophyletic group).
This study has detached from many of the previous assumptions on dinosaur phylogeny and has analysed a large number of species and many characters not included in previous investigations. This has made the resulting evolutionary tree pretty different from the ones obtained before.
RESHAPING THE TREE
Then, how does the dinosaur’s evolutionary tree stand according to this hypothesis? Well, the matter is somewhat complex, even if the different groups are still divided in two orders:
Order Saurischia which, according to this study, only includes sauropodomorphs and herrerasaurids (a group of carnivorous, non-theropod saurischians).
The new order Ornithoscelida (bird-limbed) that includes the traditional ornithischians and theropods, which are no longer saurischians.
Keeping this in mind, let’s now see the characteristics that define these two orders.
Saurischians
The order Saurischia is almost the same, except that theropods are no longer part of this group. This order presents the original saurischian hip structure, which the dinosaurs’ ancestors also had. According to this new hypothesis, herrerasaurids and sauropodomorphs are all included as saurischians.
Herrerasaurids (Herrerasauridae family) were a small group of basal saurischians that evolved towards meat-eating. That’s why for a long time it was thought that they were the sister-taxon of theropods, but it was later seen that they were found among the first saurischians. Even if they were pretty specialized, they were probably displaced by competition with other predators, appearing during the middle Triassic and becoming extinct at the end of it.
Photo by Brian Smith of a Herrerasaurus skeleton and model, from the Field Museum of Natural History of Chicago.
Herrerasaurids occupied a similar ecological niche as theropods. The new hypothesis implies that hypercarnivory (feeding exclusively on meat) evolved independently twice in dinosaurs, which makes some palaeontologist question it. Yet the herrerasaurid and theropod anatomy differed in some aspects, such as the anatomy of their hands (more generalistic in herrerasaurids) and the jaw structure.
The first sauropodomorphs were biped animals just like herrerasaurids, even if they were omnivorous. Yet, sauropodomorphs would end up becoming huge herbivorous quadrupeds with characteristic long necks.
Thecodontosaurus skeleton (by Qilong), a basal sauropodomorph and a reconstruction of Plateosaurus (from Walters, Senter & Robins) a more advanced one. Even if it cannot be appreciated in this image, sauropodomorphs would increase very their size very much during their evolution (Thecodontosaurus 2 metres, Plateosaurus up to 10 metres).
Ornithoscelidans
The new dinosaur order is Ornithoscelida, which groups theropods with ornithischians. This taxon is supported by more than twenty skeletal synapomorphies (derived characters shared by a clade), present both in basal theropods and ornithischians. Some of these characteristics include the presence of a gap between premaxillar and maxillar teeth (diastema) and the fusion of the ends of the tibia and the fibula into a tibiotarsus (even if these characteristics are only found on the most basal species).
Scheme from Baron et al. (2017) of the skulls of two basal ornithoscelidans, Eoraptor (a theropod, top) and Heterodontosaurus (an ornithischian, bottom).
Both theropods and the first ornithischians were bipedal animals. Also, the presence of heterodont teeth in the ancestral members of both groups leads us to think that the first ornithoscelidans were omnivorous, which would later specialise in feeding on meat and on plants (theropods and ornithopods respectively).
Reconstruction of the face of Daemonosaurus, one of the first theropods, by DeadMonkey8984.
A curiosity about the new classification is that accepting Ornithoscelida as a valid taxon, all feathered dinosaurs are put together into one group. Everyone knows that many theropods presented feathers (as they were the ancestors of birds) but, what most people don’t know is that feathers have also been found in some basal ornithischians and in more advanced ones too.
Reconstruction by Tom Parker of Kulindadromeus, a ornithischian which feathers have been proved to be present on most of its body.
KEEP INVESTIGATING
Then, is this hypothesis irrevocable? Well, no of course. Even if it’s pretty tempting to assume that the dinosaur’s natural history has been changed, we cannot say that from now on dinosaurs will be classified this way.
Dinosaur evolutionary tree according to Baron et al. (2017), in which we can see the different clades; Dinosauria (A), Saurischia (B) and Ornithoscelida (C).
Even if this study shows really interesting results about the origin of dinosaurs, we cannot dismiss hundreds of previous studies about this group of animals. We’ll have to remain alert to new articles that step by step will keep unveiling more information about the relationships between these Mesozoic reptiles. And that’s what’s so stimulating about biology, that there’s nothing sure! And that with new investigation techniques and new discoveries, little by little we learn more about the world around us.
Keep your mind open and keep investigating!
REFERENCES
The following sources have been consulted during the elaboration of this entry:
Durante más de 130 años los dinosaurios han sido clasificados en dos órdenes separados, los saurisquios y los ornitisquios. Pero como siempre pasa en las ciencias biológicas, toda teoría es cierta hasta que se demuestra lo contrario. Un nuevo estudio ha puesto en entredicho la clasificación clásica de los dinosaurios, eliminando y redistribuyendo algunos de los diferentes grupos de dinosaurios. Aunque esta nueva hipótesis no es segura al 100%, en esta entrada os explicaremos en que consiste este reordenamiento de los dinosaurios.
CLASIFICACIÓN TRADICIONAL DE LOS DINOSAURIOS
Desde el siglo XIX, los dinosaurios se han dividido en dos grandes órdenes basados en la estructura de su pelvis. El orden Saurischia (pelvis de lagarto) incluye a los terópodos(dinosaurios carnívoros y aves actuales) y a los sauropodomorfos(grandes herbívoros de cuello largo); el orden Ornithischia (pelvis de ave) incluye a los ornitópodos(dinosaurios herbívoros y con pico de pato), a los marginocéfalos(dinosaurios con cuernos y estructuras craneales reforzadas) y a los tireóforos(dinosaurios acorazados).
Árbol evolutivo tradicional de los dinosaurios por Zureks, con los dos tipos de pelvis dinosaurias abajo.
Aun así, esta clasificación no tiene la última palabra. La paleontología es una ciencia extremadamente volátil, ya que con cada nuevo descubrimiento se puede desmontar todo lo que se sabía hasta el momento, aunque se trate de una hipótesis centenaria. Esto es lo que ha pasado con los dinosaurios recientemente.
EL INICIO DE UNA NUEVA HIPÓTESIS
Un nuevo estudio publicado el marzo de 2017, ha hecho replantear la clasificación tradicional de los dinosaurios. Muchos estudios anteriores asumían como cierta la clasificación en Saurischia/Ornithischia tradicional y por lo tanto, los caracteres y los taxones utilizados ya iban enfocados a dicha clasificación. En cambio, este nuevo estudio ha sido pionero en varios aspectos:
Incluye un gran número de especies y taxones (muchos más que en investigaciones anteriores).
Estudios anteriores daban mucho más énfasis en dinosaurios terópodos y sauropodomorfos (saurisquios tradicionales) basales, ya que fueron de los primeros grupos en diversificarse, incluyendo pocos ornitisquios basales.
También se han incluido muchos arcosaurios dinosauromorfos (taxones no dinosaurios).
Estudios anteriores habían asumido que muchos caracteres de los ornitisquios eran simplesiomórficos (caracteres ancestrales de todos los dinosaurios) y sólo se centraban en pocas sinapomorfías (caracteres compartidos por un grupo monofilético).
Este estudio se ha desprendido de muchas de las suposiciones anteriores sobre filogenia dinosauria y ha analizado un gran número de especies y multitud de caracteres no incluidos en investigaciones anteriores. Esto ha hecho que el árbol evolutivo resultante fuese muy diferente de los que se habían obtenido hasta la fecha.
RECONSTRUYENDO EL ÁRBOL
Entonces, ¿cómo queda el árbol evolutivo de los dinosaurios según esta hipótesis? Pues el tema es un poco complicado, aunque los diferentes taxones siguen quedando divididos en dos órdenes:
El orden Saurischia que, según el estudio, sólo incluye a los sauropodomorfos y a los herrerasáuridos (grupo de saurisquios carnívoros no terópodos).
El nuevo orden Ornithoscelida (patas de ave) que incluye a los ornitisquios tradicionales y a los terópodos, que dejan de ser saurisquios.
Teniendo esto en mente, veamos ahora las características que definen a estos dos grupos.
Saurisquios
El orden Saurischia se mantiene prácticamente igual, excepto que los terópodos quedan fuera del grupo. Este orden presenta la pelvis con estructura saurisquia original, como también la presentaban los antepasados de los dinosaurios. Según la nueva hipótesis, dentro de los saurisquios se incluyen los herrerasáuridos y los sauropodomorfos.
Los herrerasáuridos (familia Herrerasauridae) fueron un pequeño grupo de saurisquios basales que evolucionaron hacia una dieta carnívora. Por eso durante mucho tiempo se pensó que eran el grupo hermano de los terópodos, pero después se vio que su lugar se encontraba entre los primeros saurisquios. Aun estando bastante especializados, probablemente la competencia con otros depredadores hizo que no durasen mucho tiempo, apareciendo a mediados del Triásico y extinguiéndose a finales de este período.
Foto de Brian Smith de un esqueleto y un modelo de Herrerasaurus del Field Museum of Natural History de Chicago.
Los herrerasáuridos ocupaban un nicho ecológico similar al de los terópodos. La nueva hipótesis implica entonces que la hipercarnivoría (alimentación exclusivamente de carne) evolucionó independientemente dos veces en los dinosaurios, cosa que hace que algunos paleontólogos la pongan en duda. Aun así, la anatomía de los herrerasáuridos y de los terópodos difería en ciertos aspectos, como la anatomía de sus manos (más generalista en los herrerasáuridos) y la estructura de la mandíbula.
Los primeros sauropodomorfos eran animales bípedos igual que los herrerasáuridos, aunque éstos eran animales omnívoros. Aun así, los sauropodomorfos acabarían convirtiéndose en enormes cuadrúpedos herbívoros con cuellos larguísimos característicos.
Esqueleto de Thecodontosaurus (por Qilong), un sauropodomorfo basal y una reconstrucción de Plateosaurus (por Walters, Senter & Robins) uno de más avanzado. Aunque en esta imagen no se aprecia, a lo largo de su evolución, los sauropodomorfos aumentarían mucho su tamaño (Thecodontosaurus 2 metros, Plateosaurus hasta 10 metros).
Ornitoscélidos
El nuevo orden de dinosaurios es Ornithoscelida, que agrupa los terópodos con los ornitisquios. Este taxón está apoyado por más de veinte sinapomorfías (caracteres derivados compartidos por un clado) esqueléticas, presentes tanto en terópodos como en ornitisquios basales. Algunas de estas características incluyen la presencia de una separación entre los dientes premaxilares y maxilares (diastema) y la fusión de los extremos de la tibia y la fíbula en un tibiotarso (aunque estos caracteres sólo se encuentran en las especies más basales).
Esquema de Baron et al. (2017) de los cráneos de dos ornitoscélidos basales, Eoraptor (un terópodo, superior) y Heterodontosaurus (un ornitisquio, inferior).
Tanto los terópodos como los primeros ornitisquios eran animales bípedos. Además, la presencia de dientes heterodontos en los miembros ancestrales de ambos grupos nos lleva a pensar que los primeros ornitoscélidos eran omnívoros, los cuáles después se especializarían en alimentarse de carne y de vegetación (terópodos y ornitisquios respectivamente).
Reconstrucción de la cara de Daemonosaurus, uno de los primeros terópodos, por DeadMonkey8984.
Una curiosidad de la nueva clasificación es que aceptando a Ornithoscelida como un taxón válido, se agrupan a todos los dinosaurios con plumas en un solo grupo. Por todos es sabido que muchos terópodos presentaban plumas (ya que son los antepasados de las aves) pero, lo que mucha gente desconoce es que la presencia de plumas también se ha demostrado en algunos ornitisquios basales y en otros más avanzados.
Reconstrucción por Tom Parker de Kulindadromeus, un ornitisquio del cual se han encontrado pruebas de que presentaba plumas en gran parte del cuerpo.
SEGUIR INVESTIGANDO
Y entonces, ¿es definitiva esta hipótesis? Pues no, por supuesto. Aunque resulta bastante atractivo decir que se ha cambiado la historia natural de los dinosaurios para siempre, no podemos asegurar que a partir de ahora los dinosaurios se clasifiquen de este modo.
Árbol evolutivo de los dinosaurios según Baron et al. (2017), donde vemos los diferente clados; Dinosauria (A), Saurischia (B) y Ornithoscelida (C).
Aunque este estudio muestre resultados interesantes sobre el origen de los dinosaurios, no se pueden ignorar los cientos de estudios anteriores que se han hecho sobre este grupo de animales. Deberemos estar atentos a nuevos artículos que poco a poco vayan desvelando más información sobre las relaciones entre estos reptiles mesozoicos. ¡Y es que esto es lo más estimulante de la biología, que no hay nada seguro! Y que con nuevas técnicas de estudio y nuevos descubrimientos, poco a poco aprendemos más sobre el mundo que nos rodea.
¡Mantened la mente abierta y seguid investigando!
REFERENCIAS
Se han consultado las siguientes fuentes durante la elaboración de esta entrada:
Durant més de 130 anys els dinosaures han estat classificats en dos ordres separats, els saurisquis i els ornitisquis. Però com sempre passa en les ciències biològiques, tota teoria és certa fins que es demostra el contrari. Un nou estudi ha posat en entredit la classificació clàssica dels dinosaures, eliminant i redistribuïnt alguns dels diferents grups de dinosaures. Tot i que aquesta nova hipòtesis no és segura al 100%, en aquesta entrada us explicarem en què consisteix aquesta reordenació dels dinosaures.
CLASSIFICACIÓ TRADICIONAL DELS DINOSAURES
Des del segle XIX, els dinosaures s’han dividit en dos grans ordres basats en la estructura de la seva pelvis. L’ordre Saurischia (pelvis de llangardaix) inclou als teròpodes(dinosaures carnívors i ocells actuals) i als sauropodomorfs(grans herbívors de coll llarg); l’ordre Ornithischia (pelvis d’ocell) inclou als ornitòpodes(dinosaures herbívors i amb bec d’ànec), als marginocèfals(dinosaures amb banyes i estructures cranials endurides) i als tireòfors(dinosaures cuirassats).
Arbre evolutiu tradicional dels dinosaures per Zureks, amb els dos tipus de pelvis dinosauries a baix.
Tanmateix, aquesta classificació no té l’última paraula. La paleontologia és una ciència extremadament volàtil, ja que amb cada nou descobriment es pot desmuntar tot el que es sabia fins al moment, encara que es tracti d’una hipòtesi centenària. Això és el que ha passat amb els dinosaures recentment.
L’INICI D’UNA NOVA HIPÒTESI
Un nou estudi publicat el març del 2017, ha fet replantejar la classificació tradicional del dinosaures. Molts estudis anteriors assumien com a certa la classificació en Saurischia/Ornithischia tradicional i per tant, els caràcters i els taxons utilitzats ja anaven enfocats en aquesta classificació. En canvi, aquest nou estudi ha estat pioner en varis aspectes:
Inclou un gran nombre d’espècies i taxons (molts més que en investigacions anteriors).
Estudis anteriors donaven molt més ènfasi en dinosaures teròpodes i sauropodomorfs (saurisquis tradicionals) basals, ja que van ser dels primers grups en diversificar-se, incloent pocs ornitisquis basals.
També s’hi han inclòs molts arcosaures dinosauromorfs (taxons no dinosaures).
Estudis anteriors havien assumit que molts caràcters dels ornitisquis eren simplesiomòrfics (caràcters ancestrals de tots els dinosaures) i només es centraven en poques sinapomorfies (caràcters compartits per un grup monofilètic).
Aquest estudi s’ha desprès de moltes de les presuposicions anteriors sobre filogènia dinosauria i ha analitzat un gran nombre de espècies i multitud de caràcters no inclosos en investigacions anteriors. Això ha fet que l’arbre evolutiu resultant fos molt diferent dels que s’havien obtingut fins llavors.
RECONSTRUÏNT L’ARBRE
Llavors, com queda l’arbre evolutiu dels dinosaures segons aquesta hipòtesi? Doncs el tema és una mica complicat, tot i que els diferents taxons segueixen quedant dividits en dos ordres:
L’ordre Saurischia que, segons l’estudi, només inclou als sauropodomorfs i als herrerasàurids (grup de saurisquis carnívors no teròpodes).
El nou ordre Ornithoscelida (potes d’ocell) que inclou als ornitisquis tradicionals i als teròpodes, que deixen de ser saurisquis.
Tenint això en ment, vegem doncs les característiques que defineixen aquests dos grups.
Saurisquis
L’ordre Saurischia es manté pràcticament igual, excepte que els teròpodes es queden fora del grup. Aquest ordre presenta la pelvis d’estructura saurisquia original, com també la presentaven els avantpassats dels dinosaures. Segons la nova hipòtesi, dins dels saurisquis s’hi inclouen els herrerasàurids i els sauropodomorfs.
Els herrerasàurids (família Herrerasauridae) van ser un petit grup de saurisquis basals que van evolucionar cap a una dieta carnívora. Per això durant un temps es pensava que eren el grup germà dels teròpodes, però després es va veure que el seu lloc es trobava entre els primers saurisquis. Tot i estar força especialitzats, probablement la competència amb altres depredadors va fer que no duressin molt de temps, apareixent a mitjans del Triàssic i extingint-se al finals d’aquest període.
Foto de Brian Smith d’un esquelet i un model de Herrerasaurus del Field Museum of Natural History de Chicago.
Els herrerasàurids ocupaven un nínxol ecològic similar al dels teròpodes. La nova hipòtesi implica doncs que la hipercanivoria (alimentació exclusivament de carn) va evolucionar independentment dos cops en els dinosaures, cosa que fa que alguns paleontòlegs la posin en dubte. Tanmateix, l’anatomia dels herrerasàurids i dels teròpodes diferia en certs aspectes, com la anatomia de les mans (més generalista en els herrerasàurids) i la estructura de la mandíbula.
Els primers sauropodomorfs eren animals bípedes igual que els herrerasàurids, tot i que aquests eren animals omnívors. Tanmateix, els sauropodomorfs acabarien convertint-se en enormes quadrúpedes herbívors amb colls llarguíssims característics.
Esquelet de Thecodontosaurus (per Qilong), un sauropodomorf basal i una reconstrucció de Plateosaurus (per Walters, Senter & Robins) un de més avançat. Tot i que en aquesta imatge no s’aprecia, al llarg de la seva evolució, els sauropodomorfs augmentarien molt la seva mida (Thecodontosaurus 2 metres, Plateosaurus fins a 10 metres).
Ornitoscèlids
El nou ordre de dinosaures és Ornithoscelida, que agrupa els teròpodes amb els ornitisquis. Aquest taxó està recolzada per més de vint sinapomorfies (caràcters derivats compartits per un clade) esquelètiques, presents tant en teròpodes com en ornitisquis basals. Algunes d’aquestes característiques inclouen la presència d’una separació entre les dents premaxil·lars i maxil·lars (diastema) i la fusió dels extrems de la tíbia i la fíbula en un tibiotars (tot i que aquests caràcters només es troben en les espècies més basals).
Esquema de Baron et al. (2017) dels cranis de dos ornitoscèlids basals, Eoraptor (un teròpode, superior) i Heterodontosaurus (un ornitisqui, inferior).
Tant els teròpodes com els primers ornitisquis eren animals bípedes. A més, la presència de dents heterodontes en els membres ancestrals d’ambdós grups fa pensar que els primers ornitoscèlids eren omnívors, els quals després s’especialitzarien en alimentar-se de carn i de vegetació (teròpodes i ornitisquis respectivament).
Reconstrucció de la cara de Daemonosaurus, un dels primers teròpodes, per DeadMonkey8984.
Una curiositat de la nova classificació és que acceptant a Ornithoscelida com un taxó vàlid, s’agrupen a tots els dinosaures amb plomes en un sol grup. Per tots és sabut que molts teròpodes presentaven plomes (ja que eren els avantpassats dels ocells) però, el que molta gent desconeix és que la presència de plomes també s’ha demostrat en alguns ornitisquis basals i en altres més avançats.
Reconstrucció per Tom Parker de Kulindadromeus, un ornitisqui del qual s’han trobat proves que presentava plomes a gran part del cos.
SEGUIR INVESTIGANT
I llavors, és definitiva aquesta hipòtesi? Doncs no, per descomptat. Tot i que resulta força atractiu dir que s’ha canviat la història natural dels dinosaures per sempre, no podem dir que a partir d’ara els dinosaures es classifiquin d’aquesta manera.
Arbre evolutiu dels dinosaures segons Baron et al. (2017), on veiem els diferents clades; Dinosauria (A), Saurischia (B) i Ornithoscelida (C).
Encara que aquest estudi mostri resultats interessants sobre l’origen dels dinosaures, no es poden ignorar els centenars d’estudis anteriors que s’han fet sobre aquest grup d’animals. Haurem d’estar atents a nous articles que poc a poc vagin desvelant més informació sobre les relacions entre aquests rèptils mesozoics. I és que això és el més estimulant de la biologia, que no hi ha res segur! I que amb noves tècniques d’estudi i nous descobriments, poc a poc aprenem més sobre el món que ens envolta.
Mantingueu la ment oberta i seguiu investigant!
REFERÈNCIES
S’han consultat les següents fonts durant l’elaboració d’aquesta entrada:
The presence of feathers is one the main characteristics of modern birds. Currently many dinosaur fossils show us that feathers appeared long before birds. Yet the feathers that those Mesozoic animals had weren’t exactly the same as the ones current birds have. The evolution of feathers was a long and gradual process, and in this entry we’ll review the most important evolutionary stages that brought those dinosaurs to develop anatomically modern feathers.
If you like this article and believe that our blog is useful, we ask for your vote in Bitácoras Awards 2015 as the best blog of Education and Science. Click the image below, enter with your Facebook or Twitter account, type the URL of the blog in the category “Educación y ciencia” and press enter. In just 30 seconds you will help us to spread more science! Please leave a comment if you have any question.
Today’s feathers
Feathers are fundamental structures for the life of modern birds. Feathers help them insulate from cold and hot weather, make them waterproof, camouflage, allow them to fly and in many species, feathers are very important in the mating rituals. In many birds, plumage allows us to differentiate between different species, telling a male and a female apart, and even allows us to know the age of an individual.
Male golden pheasant (Chrysolophus pictus) photographed at Kuala Lumpur’s Bird Park, showing us different types of feathers. Photo by Bjørn Christian Tørrissen.
Feathers are the most complex integumentary structures found in vertebrates. These are formed in the epidermis, in little follicles which produce keratin. The β-keratin of the bird’s feathers, claws and beak is much more folded than the α-keratin found in mammalian’s hair, hooves or horns, making the first a much stronger structure. Feathers are resistant and light structures, but in many birds they correspond to a third of their body weight.
Modern feathers have a central shaft divided into two parts: the proximal part which inserts to the body called the calamus, and the rachis, the distal part from which the laminar part of the feather appears. This is called the vane and is disposed on both sides of the rachis. The laminar part is made of parallel ramifications called barbs, which have ramifications called barbules which also have ramifications in the shape of small hooks called barbicels that make barbules cross-attach to each other. The superior end of the vane (pennaceous part) barbules are perfectly arranged by the barbicels, while in the inferior end (plumulaceus part) barbules lack barbicels and so they float free from each other.
According to its structure, in current birds we can find two main types of feathers:
Contour feathers: These are the feathers that make up the shape of the bird. These are long, flat feathers with a well-developed rachis and well-arranged barbs. These can be further classified into generic contour feathers, which cover the head, neck, trunk and limbs of the animal, and the flight feathers, called rectrices the ones in the tail (symmetric) and remiges the ones in the wings (asymmetric).
Down feathers: These are found forming a second layer under the contour feathers. These are feathers with a short rachis and with disordered barbs floating freely. Its main function is to thermally insulate the bird. Natal down feathers covering most bird hatchlings in some time of their lives are called “neossoptilus”.
Barn owl hatchling (Tyto alba) covered in down feathers. Photo by Maxgreen.
Apart from these two types, there are other kinds of feathers in birds, such as the semiplumes (with an intermediate structure between contour and down feathers) and the bristles and filoplumes (with few barbs and mainly with a sensory function).
Different types of feathers we can find on modern birds, drawings by Osado. From left to right: Rectrix (tail), remex (wing), generic contour feather, semiplume, down feather, bristle and filoplume.
Origin and evolution of feathers
Probably dinosaurs develped the first feathers as a system to avoid the loss of body heat. Having a covering feathers, a layer of warm air becomes trapped around the animal, making its body temperature more stable. That’s why some scientists think that many dinosaur species had an almost endothermic metabolism (mesothermy), with high and constant body temperature. Nevertheless, primitive feathers or “protofeathers” were very different from modern feathers.
Reconstruction of Deinonynchus by Stephen Czerkas, at the Natural History Museum of Vienna. Photo by Domser.
As we will now see, protofeathers went through different evolutionary stages before becoming modern feathers. Even if here we present you these stages linearly, it doesn’t mean that when a new kind of protofeather appeared the previous one disappeared. Just like modern birds sport different kinds of feathers, many dinosaurs presented different combinations of protofeathers, which only represented different levels of specialization.
Stage 1: A single filament
Drawing about the origin and formation of the first protofeathers. Extracted from Prum & Brush (2002).
The first known protofeathers were nothing more than a cylindrical hollow spine-like filament, which formed on a follicle’s collar. Even though feathers and protofeathers are typically exclusive characteristics of theropods, this first protofeathers have also been found in two groups of non-theropod dinosaurs. These are the Heterodontosauridae and Psittacosauridae families, many species of which had spines homologous to stage 1 protofeathers which probably also served to retain body heat.
Reconstruction of a heterodontosaurid named Fruitadens. Drawing by Smokeybjb.
In theropods, feathers appeared in a group named Coelurosauria, which includes animals like the tyrannosaur, the velociraptor and modern birds. The oldest feathered coelurosaur known is Sciurumimus, which literally means “squirrel mimic”. This fossil got its name for its fully feathered tail, covered in filamentous protofeathers similar to a squirrel’s.
Reconstruction of a juvenile Sciurumimus based on the skeleton found in Bavaria. Drawing by Franz Joseph.
Stage 2: A plumulaceous protofeather
Second stage in the evolution of feathers, in which a division in the follicle produces various barbs with a single origin. Extracted from Prum & Brush (2002).
The next step on the evolution of feathers was the division of the cellular collar of the follicle, which brought the branching of the filament. The result is a plumulaceous protofeather with unbranched barbs originating in a calamus. Stage 2 protofeather are similar to down feathers of current birds and have been found in a wide variety of theropod fossils.
These protofeathers provided a better insulation, helping the animal to keep its body heat. It is also believed that it’s likely that the smallest dinosaurs were more fully covered in protofeathers, since smaller animals loose heat faster than bigger animals and so, they need more mechanisms to retain body heat. Bigger coelurosaur species like Tyrannosaurus may have lost their protofeathers much like modern elephants have lost almost all their body hair. Yet, it is possible that some species presented protofeathers after birth and during the first stages of life, and after growing up they would either loose them or only present them on some body parts.
Reconstruction of a juvenile Juravenator in which we can appreciate how it was covered both with protofeathers and scales. Drawing by Tom Parker.
Yet in a Chinese paleontological site, the two biggest feathered dinosaurs known were discovered. The first to be discovered was Beipiaosaurus, a strange looking coelurosaur of about 3 metres long with long claws, which presented protofeathers both filamentous (stage 1) and plumulaceous (stage 2). This species shared its habitat with Yutyrannus, a 9 metre-long animal up to 1400 kilos of weight, which had almost all its body covered in plumulaceous protofeathers. These two animals probably lived in a humid and cold environment, and their coat of protofeathers helped them to keep their warmth when temperatures would fall.
Reconstruction of four Yutyrannus and a pair of Beipiaosaurus on their habitat. Drawing by Brian Choo.
Stage 3: Fusion and branching
Drawing of the evolution of feathers from stage 1 to 3. Extracted from Sues (2001).
The third stage in the evolution of feathers gave rise to a protofeather with a central rachis made from the fusion of some barbs (3a) and a protofeather with barbules branching from the main barbs (3b). The combination of these two characters produced a pennaceous, vaned protofeather similar to the ones found in modern birds but less firm, as it lacked the hooked barbicels of modern feathers.
Fossils of stage 3a protofeathers where we can see a central rachis from which various barbs extend. Extracted from Perrichot (2008).
Stage 4: Hooks to maintain order
Modified drawing from Prum & Brush (2002) of the apparition of hooks on the barbules of the stage 4.
It is in this stage where we can start talking about present day feathers. The stage 3 structure with a rachis, barbs and barbules, developed small hooks on the barbules which made them cross-attach and keep the vane together. These feathers were like the ones found in modern birds, the contour feathers, which present a central shaft and a symmetric vane.
Reconstruction of the troodontid called Anchiornis, where the wide cover of feathers it presented can be seen. Drawing by Matt Martyniuk.
These feathers are found in many different dinosaurs, many of which had begun to develop adaptations for flight, or at least gliding. Despite this, we can also find them in typically running dinosaurs like Velociraptor, a terrestrial predator about the size of a turkey, with a long mouth and a sickle-shaped claw on its hind legs. This claw is thought to be used mainly to kill their prey, but some scientists think that they used their claw to climb trees and ambush their prey from above. Maybe their feathers served them to make more controlled leaps when they fell on their victims.
Drawing of a velociraptor attacking an oviraptotosaur. Drawing by Durbed.
These feathers are also found in the oviraptorosaurs, a coelurosaurian group with beak and few or no teeth. Even if they couldn’t fly, they probably used their arm feathers to incubate their eggs (like the Avimimus genre) and the ones on the tail for display and communication with other members of their species (like the Caudipteryx, and Nomingia genera).
Reconstruction of the oviraptorosaurian Nomingia, in which we can see the fan of feathers on its tail. Drawing by Smokeybjb.
Other dinosaurs like Scansoriopteryx had an arboreal lifestyle, and the feathers on its arms allowed it to glide from one tree to the other both to hunt and to escape predators. A relative of this animal called Epidexipteryx, even though not having feathers on its arms (as far as is known) presented long vaned feathers on the tail, probably to send visual signals to other members of its species.
Reconstruction of Epidexipteryx in which the long vaned tail feathers can be appreciated. Reconstruction by Nobu Tamura.
Stage 5: Asymmetry brings flight
Drawings and fossils of all the different stages of the evolution of feathers. Extracted from McKellar et al (2011).
Finally the last stage in the evolution of feathers is the appearance of asymmetric feathers, with a displaced rachis making one half of the vane wider than the other. These feathers are the remiges found on the wings of birds, which not only increase drag during gliding, but also allow the animal to leave the ground and fly.
Photo of a Willow tit (Poecile montanus) taking flight, where we can perfectly see the asymmetric remiges on its wings. Photo by David Mark.
Even if it’s generally assumed that apart from birds no other dinosaur group accomplished powered flight, there’s one group which got really close. The microraptorians were a group of small feathered dinosaurs characterized by presenting flight feathers, not only on their front limbs but also on their hind limbs. The most famous of them, Microraptor, had asymmetrical flight feathers on its arms, legs and, unlike modern birds, on its tail.
Drawing of the silhouette of Microraptor gliding. Extracted from Xu et al.
Even if it’s usually considered a glider, some authors argue that possibly Microraptor was capacitated to fly. Some skeletal characteristics indicate that some microraptorians may have been better suited for flight than Archaeopteryx, the ancestor of modern birds. For example, Microraptor presented a fused more developed sternum than Archaeopteryx, which would have made for a major anchor point for the flight muscles.
Nevertheless, Archaeopteryx is still considered the nearest to the ancestor of modern birds that, even if it wasn’t a great flier, it already had the different kinds of feathers found on current birds. Probably many more dinosaurs were covered with feathers or protofeathers, but in this entry we have only seen the species which show irrefutable evidence of having them. As we have seen, the road to reach modern feathers was long and gave rise to a wide diversity of dinosaur species, but after a meteorite practically extinguished life on Earth 65 million years ago, only one group of feathered dinosaurs survived and thrived.
Fossil of Archaeopteryx lithographica from the late Jurassic found at southern Germany. Photo by James L. Amos.
Did you enjoyed it? Remember to assess our work in Bitácoras Awards voting for us as the best blog of Education! Thanks a lot.
References
The next sources have been consulted during the elaboration of this entry:
La presencia de plumas es una de las características más destacables de los pájaros actuales. Actualmente son muchos los fósiles de dinosaurios que nos indican que las plumas aparecieron mucho antes que las aves modernas. Aun así, las plumas que presentaban estos animales mesozoicos no eran exactamente iguales a las que tienen los pájaros de hoy en día. La evolución de las plumas fue un proceso largo y gradual, y en esta entrada intentaremos hacer un repaso de los principales estadios evolutivos que llevaron a estos dinosaurios a desarrollar las plumas actuales.
Si te gusta este artículo y crees que nuestro blog es útil, te pedimos por favor tu voto en los Premios Bitácoras 2015 como el mejor blog de Educación y Ciencia. Pincha en la imagen siguiente, entra con tu cuenta de Facebook o Twitter, escribe la URL del blog en la categoría “Educación y Ciencia” y pulsa enter. ¡en sólo 30 segundos nos ayudarás a divulgar más ciencia! Por favor, deja un comentario para cualquier duda.
Plumas actuales
Las plumas son estructuras fundamentales para la vida de los pájaros actuales. Les proporcionan aislamiento contra el frío, el calor y el agua, camuflaje, les permiten el vuelo y en muchas especies son importantísimas en los rituales de apareamiento. En muchas aves, el plumaje permite distinguir entre especies diferentes, entre machos y hembras, e incluso hasta nos permite saber la edad de un individuo.
Macho de faisán dorado (Chrysolophus pictus) fotografiado en el Bird Park de Kuala Lumpur mostrándonos diversas morfologías de plumas. Foto de Bjørn Christian Tørrissen.
Las plumas son las estructuras tegumentarias más complejas de todas las que encontramos entre los vertebrados. Éstas se forman en la epidermis, en pequeños folículos que producen queratina. La β-queratina de las plumas, las garras y el pico de las aves se encuentra mucho más replegada que la α-queratina del pelo, las pezuñas y los cuernos de los mamíferos, haciendo que sea mucho más fuerte la estructura de la primera. Las plumas son estructuras resistentes y ligeras, aunque en muchos pájaros corresponden a un tercio de su peso corporal.
Las plumas actuales presentan un eje dividido en dos partes: el cálamo o cañón, la parte más proximal y que se inserta en el cuerpo, y el raquis, la parte más distal y de donde sale la parte laminar de la pluma. Ésta se llama vexilo o estandarte y está dispuesta a ambos lados del raquis. La parte laminar está compuesta por ramificaciones paralelas llamadas barbas, las cuáles a su vez se ramifican en barbillas o bárbulas, las cuáles también se ramifican en pequeños ganchos que hacen que las bárbulas se enganchen entre ellas. En la parte superior del vexilo (parte plumácea) las barbas quedan perfectamente ordenadas por el sistema de ganchos, mientras que en la parte inferior (parte plumosa) las barbas están separadas y desordenadas, ya que presentan pocos ganchos.
Según su estructura, en los pájaros actuales encontramos dos tipos principales de plumas:
Plumas de contorno: Son las plumas que dan forma al pájaro. Son plumas largas y planas, con el raquis desarrollado y barbas ordenadas. Éstas se pueden clasificar en las plumas genéricas de contorno, que cubren cabeza, cuello, tronco y miembros del animal, y las plumas de vuelo, llamadas timoneras o rectrices las de la cola (simétricas) y remeras o rémiges las de las alas (asimétricas).
Plumón: Éstas se encuentran formando una segunda capa debajo de las plumas de contorno. Son plumas con un raquis corto y con las barbas desordenadas y separadas entre ellas. Su función principal es la de aislar térmicamente al pájaro. El plumón natal que recubre los polluelos de la mayoría de aves en algún momento de su desarrollo se llama “neossoptilus”.
Polluelo de lechuza (Tyto alba) cubierto de plumón. Foto de Maxgreen.
Aparte de estos dos tipos, en los pájaros hay otras clases de plumas, como las semiplumas (con una estructura intermedia entre las de contorno y el plumón) y las cerdas y filoplumas (con pocas barbas y función principalmente sensorial).
Dibujos esquemáticos de los diferentes tipos de plumas que encontramos en las aves actuales, realizados por Osado. De izquierda a derecha: Rectriz (cola), rémige (ala), pluma genérica de contorno, semipluma, plumón, cerda y filopluma.
Origen y evolución de las plumas
Probablemente los dinosaurios desarrollaron las primeras plumas como sistema para evitar la pérdida de calor corporal. Teniendo una cobertura de plumas se crea una capa de aire caliente alrededor del animal haciendo que su temperatura sea más estable. Por eso se cree que muchos dinosaurios tenían un metabolismo casi endotérmico (mesotermia) con una temperatura alta y constante. Aun así, las plumas primitivas o “protoplumas” eran muy diferentes de las plumas actuales.
Reconstrucción de Deinonynchus hecha por Stephen Czerkas, en el Museo de Historia Natural de Viena. Foto de Domser.
Como veremos a continuación, las protoplumas pasaron por diferentes estadios evolutivos hasta llegar a las plumas actuales. Aunque aquí os presentamos estos estadios de forma lineal, no significa que al aparecer un nuevo tipo de protopluma desapareciera el anterior. Igual que los pájaros actuales presentan diferentes tipos de plumas, muchos dinosaurios presentaban diferentes combinaciones de protoplumas, las cuales solo representan diferentes niveles de especialización evolutiva.
Estadio 1: Un único filamento
Esquema sobre el origen y formación de las primeras protoplumas. Extret de Prum & Brush (2002).
Las primeras protoplumas de las que se tiene constancia no eran más que unos tubos cilíndricos huecos, más parecidos a espinas que a plumas, que provenían del collar de un folículo. Aunque las plumas y protoplumas son características típicas de los terópodos, estas primeras protoplumas también se han encontrado en dos grupos de dinosaurios no terópodos. Éstos son las familias Heterodontosauridae y Psittacosauridae muchos miembros de las cuáles presentaban espinas homólogas a las protoplumas de los terópodos que seguramente también tenían la función de retener el calor corporal.
Reconstrucción del heterodontosáurido llamado Fruitadens. Dibujo de Smokeybjb.
En los terópodos, las plumas aparecieron en un grupo llamado Coelurosauria, que incluye a animales como el tiranosaurio, el velociraptor y las aves actuales. El coelurosaurio con protoplumas más antiguo del que se tiene constancia es Sciurumimus, literalmente “imitador de ardillas”. Se llamó así a este fósil por su cola cubierta de protoplumas filamentosas, parecida a la cola de una ardilla.
Reconstrucción de un juvenil de Sciurumimus basado en un esqueleto encontrado en Baviera. Dibujo de Franz Joseph.
Estadio 2: Un abanico de barbas
Segundo estadio de la evolución de las plumas, donde una división del folículo hace que se formen varias barbas con un mismo origen. Extraído de Prum & Brush (2002).
El siguiente paso en la evolución de las plumas fue la división del collar celular del folículo para hace que el filamento resultante se ramifique. El resultado es un abanico de barbas que se extienden desde el cálamo. Las protoplumas de este estadio se parecen al plumón actual de los pájaros (protoplumas plumosas) y se han encontrado en una gran variedad de fósiles de terópodos.
Estas protoplumas proporcionaban un mayor aislamiento, ayudando al animal a conservar su temperatura corporal. Se cree también, que es más probable que los dinosaurios con el cuerpo más recubiertos de protoplumas fueran los más pequeños, ya que los animales pequeños pierdan calor más rápidamente que los animales más grandes y por los tanto necesitan más mecanismos para retener el calor corporal. Las especies de coelurosaurios más grandes como Tyrannosaurus quizás habrían perdido las protoplumas de manera similar a como los elefantes actuales han perdido prácticamente todo el pelo del cuerpo. Aun así, es posible que algunas especies presentara protoplumas al nacer y durante las primeras etapas de su vida, y que al crecer o las perdiesen o solo las presentasen en algunas partes del cuerpo.
Reconstrucción de un juvenil de Juravenator donde vemos que presentaba tanto protoplumas como escamas. Dibujo de Tom Parker.
Aun así, en un yacimiento de China, se encontraron los restos de los dos dinosaurios con plumas más grandes de los que se tiene constancia. El primero que se descubrió fue Beipiaosaurus, un extraño coleurosaurio de 3 metros de largo con largas uñas, que presentaba protoplumas tanto filamentosas (estadio 1) como plumosas (estadio 2). Éste compartía su hábitat con Yutyrannus, un animal de 9 metros de largo y 1400 kilos de peso que presentaba prácticamente todo el cuerpo cubierto de protoplumas plumosas. Estos dos animales probablemente vivían en una ambiente frío y húmedo, y su capa de protoplumas les ayudaba a conservar su calor cuando bajaban las temperaturas.
Reconstrucción de cuatro Yutyrannus y un par de Beipiaosaurus en su hábitat. Dibujo de Brian Choo.
Estadio 3: Fusión y ramificación
Esquema de la evolución de las plumas del estadio 1 al 3. Extraído de Sues (2001).
El tercer estadio en la evolución de las plumas es el que dio lugar a una protopluma con un raquis central formado por la fusión de algunas barbas (3a), y una en la que aparecieron bárbulas que surgían de las barbas (3b). La combinación de estos dos caracteres dio una protopluma plumácea, bipenada, parecida a las plumas actuales de los pájaros, aunque al no tener ganchos para unir las bárbulas entre ellas no tendrían la solidez de las plumas modernas.
Fósiles de protoplumas del estadio 3a donde se ve un raquis central de donde salen varias barbas. Extraído de Perrichot (2008).
Estadio 4: Ganchos para mantener el orden
Esquema modificado de Prum & Brush (2002) sobre la aparición de los ganchos entre las bárbulas del estadio 4.
En este estadio es donde podemos empezar a hablar de plumas propiamente dichas. La estructura con raquis, barbas y bárbulas del estadio 3, desarrolló unos pequeños ganchos en las bárbulas que hacían que éstas se engancharan y mantuvieran el vexilo unido y con las barbas ordenadas. Estas plumas son las que presentan los pájaros actuales, las llamadas plumas de contorno, las cuales presentan un raquis central y un vexilo simétrico.
Reconstrucción del troodóntido Anchiornis, donde se aprecia la amplia cobertura de plumas que presentaba. Dibujo de Matt Martyniuk.
Estas plumas las encontramos en diferentes dinosaurios, muchos de los cuales ya empezaban a desarrollar adaptaciones para volar, o al menos planear. Aun así, también se encuentran en dinosaurios típicamente corredores como el Velociraptor, un depredador terrestre del tamaño de un pavo, con un hocico largo y una garra con forma de hoz en las patas traseras. La garra se cree que servía para matar a las presas, aunque algunos científicos creen que utilizaban la garra para subirse a los árboles y emboscar las presas desde arriba. Quizás las plumas le habrían servido para controlar el salto cuando se abalanzaba sobre sus víctimas.
Dibujo de un velociraptor atacando a un oviraptorosaurio. Dibujo de Durbed.
Estas plumas también las encotramos en los oviraptorosaurios, un grupo de coelurosaurios con pico y pocos o ningún diente. Aunque no podían volar, este grupo probablemente utilizaba las plumas de los brazos para incubar los huevos (como en el género Avimimus) y las de la cola para exhibirse y comunicarse con sus congéneres (como los géneros Caudipteryx, i Nomingia).
Reconstrucción del oviraptorosaurio Nomingia, donde se ve el abanico de plumas en la cola. Dibujo de Smokeybjb.
Otros dinosaurios como Scansoriopteryx tenían un estilo de vida arborícola, y las plumas de los brazos le servían para planear de árbol a árbol, tanto para cazar como para huir de sus depredadores. Un pariente de éste, el Epidexipteryx aunque no presentaba plumas en los brazos (que se sepa), presentaba largas plumas penadas en la cola, probablemente para enviar señales visuales a otros miembros de su especie.
Reconstrucción de Epidexipteryx donde se observan las largas plumas penadas de la cola. Reconstrucción de Nobu Tamura.
Estadio 5: La asimetría permite el vuelo
Esquema y fósiles de todos los pasos en la evolución de las plumas. Extraído de McKellar et al (2011).
Finalmente el último estadio en la evolución de las plumas es la aparición de plumas asimétricas, con el raquis desplazado haciendo que una parte del vexilo sea más ancha que la otra. Estas plumas son las remeras o rémiges de las alas de los pájaros, las cuáles no solo crean resistencia al aire en el planeo, sino que hacen que el animal pueda elevarse del suelo y volar.
Foto de carbonero montano (Poecile montanus) alzando el vuelo, donde se ven perfectamente las rémiges asimétricas de las alas. Foto de David Mark.
Aunque se cree que aparte de los pájaros ningún grupo de dinosaurios consiguió volar realmente, hay un grupo que se acercó mucho. Los llamados microraptores eran un grupo de dinosaurios emplumados que se caracterizaban por presentar plumas de vuelo, no sólo en las patas delanteras, sino también en las traseras. Microraptor, el más famoso de éstos, presentaba plumas de vuelo asimétricas en los brazos, las piernas y, a diferencia de los pájaros actuales, también en la cola.
Dibujo de la silueta de Microraptor planeando. Extraído de Xu et al.
Aunque generalmente se le considera un planeador, algunos autores argumentan que posiblemente Microraptor hubiese estado capacitado para volar. Algunas características esqueléticas indican que algunos microraptores podrían haber estado mejor adaptados al vuelo que Archaeopteryx, el antepasado de las aves modernas. Por ejemplo, Microraptor presentaba un esternón fusionado más desarrollado que Archaeopteryx, cosa que hacía que tuviese un mayor punto de inserción para los músculos de vuelo.
Aun así, el más cercano al antepasado de los pájaros actuales fue Archaeopteryx, el cuál, aún no siendo un gran volador, ya presentaba las plumas como las de las aves modernas. Probablemente muchos más dinosaurios estaban cubiertos de plumas o protoplumas, pero en esta entrada sólo hemos mencionado las especies de las que se tienen pruebas irrefutables de que las presentaban. Como hemos visto, el camino hasta llegar a las plumas actuales fue largo y dio lugar a una gran diversidad de especies de dinosaurios, pero después de que un meteorito extinguiera prácticamente todoa la vida de la Tierra hace 65 millones de años, sólo un grupo de dinosaurios emplumados sobrevivió y prosperó.
Fósil de Archaeopteryx lithographica de finales del Jurásico encontrado en el sur de Alemania. Foto de James L. Amos.
¿Te ha gustado? ¡Acuérdate de valorar nuestro trabajo votándonos en los Premios Bitácoras como mejor blog de Educación y Ciencia! Muchísimas gracias.
Referencias
Para la elaboración de esta entrada se han utilizado las siguientes referencias:
La presència de plomes és una de les característiques més destacables dels ocells actuals. Actualment són molts els fòssils de dinosaures que ens indiquen que les plomes van aparèixer molt abans que els ocells moderns. Tanmateix, les plomes que presentaven aquests animals mesozoics no eren exactament iguals a les que tenen els ocells d’avui en dia. L’evolució de les plomes va ser un procés llarg i gradual, i en aquesta entrada intentarem fer un repàs dels principals estadis evolutius que van portar a aquests dinosaures a desenvolupar les plomes actuals.
Si t’agrada aquest article i creus que el nostre blog és útil, et demanem per favor el teu vot en els Premis Bitácoras 2015 com el millor bloc d’Educació i Ciència. Clica la imatge següent, entra amb el teu compte de Facebook o Twitter, escriu la URL del blog a la categoria “Educació i Ciència” i prem enter. En només 30 segons ens ajudaràs a divulgar més ciència! Si us plau, deixa un comentari per a qualsevol dubte.
Plomes actuals
Les plomes són estructures fonamentals per a la vida dels ocells actuals. Els proporcionen aïllament contra el fred, la calor i l’aigua, camuflatge, els permeten el vol i en moltes espècies són importantíssimes en els rituals d’aparellament. En molts ocells, el plomatge permet diferenciar entre diferents espècies, entre mascles i femelles, i fins i tot ens permet saber l’edat d’un individu.
Mascle de faisà daurat (Chrysolophus pictus) fotografiat al Bird Park de Kuala Lumpur mostrant-nos diverses morfologies de plomes. Foto de Bjørn Christian Tørrissen.
Les plomes són les estructures tegumentàries més complexes de totes les que trobem entre els vertebrats. Aquestes es formen a l’epidermis, en petits fol·licles que produeixen queratina. La β-queratina de les plomes, les urpes i el bec dels ocells es troba molt més replegada que la α-queratina del pèl, les peülles i les banyes dels mamífers, fent que sigui molt més forta l’estructura de la primera. Les plomes són estructures resistents i lleugeres, tot i que en molts ocells corresponen a un terç del seu pes corporal.
Les plomes actuals presenten un eix dividit en dues parts: el càlem o canó, la part més proximal i que s’inserta al cos, i el raquis, la part més distal i d’ón surt la part laminar de la ploma. Aquesta s’anomena vexil o estandart i està disposada a ambdós costats del raquis. La part laminar està composta per ramificacions paralel·les anomenades barbes, les quals alhora es ramifiquen en barbetes o bàrbules, les quals també es ramifiquen en petits ganxos que fan que les bàrbules s’enganxin entre elles. A la part superior del vexil (part plomàcia) les barbes queden perfectament ordenades pel sistema de ganxos, mentre que a la part inferior (part plumosa) les barbes estan separades i desordenades, ja que presenten pocs ganxos.
Segons la seva estructura, en els ocells actuals hi trobem dos tipus principals de plomes:
Plomes de contorn: Son les plomes que dónen forma a l’ocell. Son plomes llargues i planes, amb el raquis desenvolupat i barbes ordenades. Aquestes es poden classificar en les plomes genèriques de contorn, que recobreixen el cap, coll, tronc i membres de l’animal, i les plomes de vol, anomenades timoneres o rectrius les de la cua (simètriques) i remera o rèmige les de les ales (asimètriques).
Plomissol: Aquestes es troben formant una segona capa sota les plomes de contorn. Són plomes amb raquis curt i amb les barbes desordenades i separades entre elles. La seva funció principal és la d’aïllar tèrmicament a l’ocell. El plomissol natal que recobreix els polls de la majoria d’aus en algún moment del seu desenvolupament s’anomena “neossoptilus”.
Poll d’òliba (Tyto alba) cobert de plomissol. Foto de Maxgreen.
Apart d’aquests dos tipus, en els ocells hi ha altres classes de plomes, com ara les semiplomes (amb una estructura intermitja entre les de contorn i el plomissol) i les cerdes i filoplomes (amb poques barbes i funció principalment sensorial).
Dibuixos esquemàtics dels diferents tipus de plomes que trobem en els ocells actuals, fets per Osado. D’esquerra a dreta: Rectriu (cua), rèmige (ala), ploma genèrica de contorn, semiploma, plomissol, cerda i filoploma.
Origen i evolució de les plomes
Probablement els dinosaures van desenvolupar les primeres plomes com un sistema per evitar la pèrdua d’escalfor corporal. Tenint una cobertura de plomes es crea una capa d’aire calent al voltant de l’animal fent que la seva temperatura sigui més estable. Per això es creu que molts dinosaures tenien un metabolisme quasi endotèrmic (mesotermia) amb una temperatura alta i constant. Tot i així, les plomes primitives o “protoplomes” eren molt diferents de les plomes actuals.
Reconstrucció de Deinonynchus feta per Stephen Czerkas, al Museu d’Història Natural de Viena. Foto de Domser.
Com veurem a continuació, les protoplomes van passar per diferents estadis evolutius fins a arribar a les plomes actuals. Tot i que aquí us presentem aquests estadis de forma linial, no vol dir que al aparèixer un nou tipus de protoploma desaparegués l’anterior. Igual que els ocells actuals presenten diferents tipus de plomes, molts dinosaures presentaven diferents combinacions de protoplomes, les quals només representen diferents nivells d’especialització evolutiva.
Estadi 1: Un únic filament
Esquema sobre l’origen i formació de les primeres protoplomes. Extret de Prum & Brush (2002).
Les primeres protoplomes de les que es té constància no eren res més que uns tubs cilíndrics buits, més semblants a espines que a plomes, que provenien del collar d’un fol·licle. Tot i que les plomes i protoplomes són característiques típiques dels teròpodes, aquestes primeres protoplomes també s’han trobat en dos grups de dinosaures no teròpodes. Aquests són les famílies Heterodontosauridae i Psittacosauridae molts membres de les quals presentaven espines homòlogues a les protoplomes dels teròpodes que segurament també tenien la funció de retindre l’escalfor corporal.
Reconstrucció de l’heterodontosàurid anomenat Fruitadens. Dibuix de Smokeybjb.
En els teròpodes, les plomes van apareixer en un grup anomenat Coelurosauria, que inclou a animals com el tiranosaure, el velociraptor i els ocells actuals. El coelurosaure amb protoplomes més antic del que es té constància és Sciurumimus, literalment “imitador d’esquirols”. S’anomenà així a aquest fòssil per la seva cua plena de protoplomes filamentoses, semblant a la cua d’un esquirol.
Reconstrucció d’un juvenil de Sciurumimus basat en un esquelet trobat a Baviera. Dibuix de Franz Joseph.
Estadi 2: Un plomall de barbes
Segon estadi de l’evolució de les plomes, on una divisió del fol·licle fa que es formin vàries barbes amb un mateix origen. Extret de Prum & Brush (2002).
El següent pas en l’evolució de les plomes fou la divisió del collar cel·lular del fol·licle per a fer que el filament resultant es ramifiqui. El resultat és un plomall de barbes que s’extenen des del càlem. Les protoplomes d’aquest estadi s’assemblen al plomissol actual dels ocells (protoplomes plumoses), i s’han trobat en una gran varietat de fòssils de teròpodes.
Aquestes protoplomes proporcionaven un major aïllament, ajudant a l’animal a conservar la seva temperatura corporal. Es creu també, que és més probable que els dinosaures amb el cos més recobert de protoplomes fóssin els més petits, ja que els animals petits perden escalfor més ràpidament que els animals més grans i per tant necessiten més mecanismes per a retindre l’escalfor corporal. Les espècies de coelurosaures més grans com Tyrannosaurus potser haurien perdut les protoplomes de manera similar a com els elefants actuals han perdut pràcticament tot el pèl del cos. Tot i així, és possible que algunes espècies presentessin protoplomes al néixer i durant les primeres etapes de la seva vida, i que al créixer o les perdéssin o només les presentéssin en algunes parts del cos.
Reconstrucció d’un juvenil de Juravenator on veiem que presentava tant protoplomes com escames. Dibuix de Tom Parker.
Tanmateix en un jaciment de la Xina, s’hi van trobar les restes dels dos dinosaures amb plomes més grans dels que es té constància. El primer que es va descobrir fou Beipiaosaurus, un estrany coleurosaure de 3 metres de llarg amb llargues urpes, que presentava protplomes tant filamentoses (estadi 1) com plumoses (estadi 2). Aquest compartia l’hàbitat amb Yutyrannus, un animal de 9 metres de llarg i 1400 kilos de pes que presentava pràcticament tot el cos cobert de protoplomes plumoses. Aquests dos animals probablement vivien en un ambient fred i humit, i la seva capa de protoplomes els ajudava a conservar l’escalfor quan baixaven les temperatures.
Reconstrucció de quatre Yutyrannus i un parell de Beipiaosaurus en el seu hàbitat. Dibuix de Brian Choo.
Estadi 3: Fusió i ramificació
Esquema de l’evolució de les plomes de l’estadi 1 al 3. Extret de Sues (2001).
El tercer estadi en l’evolució de les plomes és el que va donar una protoploma amb un raquis central format per la fusió d’algunes barbes (3a), i una en la que aparegueren bàrbules que sorgien de les barbes (3b). La combinació d’aquests dos caràcters donà una protoploma plumàcia, bipennada, semblant a les plomes actuals dels ocells tot i que, al no tindre ganxos per a unir les bàrbules entre elles no tindrien la solidesa de les plomes modernes.
Fòssils de protoplomes de l’estadi 3a on es veu un raquis central d’on surten vàries barbes. Extret de Perrichot (2008).
Estadi 4: Ganxos per mantindre l’ordre
Esquema modificat de Plum & Brush (2002) de l’aparició dels ganxos entre les bàrbules de l’estadi 4.
En aquest estadi és on podem començar a parlar de plomes pròpiament dites. La estructura amb raquis, barbes i bàrbules de l’estadi 3, va desenvolupar uns petits ganxos a les bàrbules que feien que aquestes s’enganxéssin i mantinguéssin el vexil unit i amb les barbes ordenades. Aquestes plomes són les que presenten els ocells actuals, les anomenades plomes de contorn, les quals presenten un raquis central i un vexil simètric.
Reconstrucció del troodòntid Anchiornis, on s’aprecia la àmplia cobertura de plomes que presentava. Dibuix de Matt Martyniuk.
Aquestes plomes les trobem en diferents dinosaures, molts dels quals ja començaven a desenvolupar adaptacions per a volar, o almenys planar. Tot i així, també es troben en dinosaures típicament corredors com el Velociraptor, un depredador terrestre de la mida d’un gall d’indi amb un musell llarg i una urpa amb forma de falç a les potes del darrere. L’urpa es creu que servia per a matar a les preses, tot i que alguns científics creuen que feien servir l’urpa per a enfilar-se als arbres i emboscar les preses des de dalt. Potser les plomes li haurien servit per a controlar el salt quan s’avalançava sobre les seves víctimes.
Reconstrucció d’un velociraptor atacant a un oviraptorosaure. Dibuix de Durbed.
Aquestes plomes també les trobem en els oviraptorosaures, un grup de coelurosaures amb bec i poques o cap dent. Tot i que no podien volar, aquest grup probablement feia servir les plomes dels braços per a incubar els ous (com en el gènere Avimimus) i les de la cua per a exhibir-se i comunicarse amb els seus congèneres (com els gèneres Caudipteryx, i Nomingia).
Reconstrucció de l’oviraptorosaure Nomingia, on es veu el ventall de plomes a la cua. Dibuix de Smokeybjb.
Altres dinosaures com Scansoriopteryx tenien un estil de vida arborícola, i les plomes dels braços li servien per planar d’arbre en arbre, tant per caçar com per fugir dels depredadors. Un parent d’aquest, el Epidexipteryx tot i no presentar plomes als braços (que se sàpiga) presentava llargues plomes pennades a la cua, probablement per enviar senyals visuals a altres membres de la seva espècie.
Reconstrucció de Epidexipteryx on s’observa les llargues plomes pennades de la cua. Reconstrucció de Nobu Tamura.
Estadi 5: L’asimetria permet el vol
Esquema i fòssils de tots els passos en l’evolució de les plomes. Extret de McKellar et al (2011).
Finalment l’últim estadi en l’evolució de les plomes és l’aparició de plomes asimètriques, amb el raquis desplaçat fent que una part del vexil sigui més ampla que l’altra. Aquestes plomes són les plomes remeres o rèmiges de les ales dels ocells, les quals no només a fer resistència a l’aire en el planeig, sinó que fan que l’animal pugui elevar-se de terra i volar.
Foto de mallerenga capnegre (Poecile montanus) alçant el vol, on es veuen perfectament les rèmiges asimètriques de les ales. Foto de David Mark.
Tot i que es creu que apart dels ocells cap grup de dinosaures va aconseguir volar realment, hi ha un grup que s’hi va apropar molt. Els anomenat microraptors eren un grup de dinosaures emplomallats que es caracteritzaven per presentar plomes de vol, no només a les potes del davant sinó també a les del darrere. Microraptor, el més famós d’aquests, presentava plomes de vol asimètriques als braços, les cames i, a diferència dels ocells actuals, també a la cua.
Dibuix de la silueta de Microraptor planant. Extret de Xu et al.
Tot i que generalment se’l considera un planador, alguns autors argumenten que possiblement Microraptor hagués estat capacitat per a volar. Algunes característiques esquelètiques indiquen que alguns microraptors podrien haver estat millor adaptats al vol que Archaeopteryx, l’avantpassat dels ocells moderns. Per exemple, Microraptor presentava un esternó fusionat més desenvolupat que Archaeopteryx, cosa que feia que tingués un major punt d’inserció per als músculs de vol.
Tanmateix, el més proper a l’avantpassat dels ocells actuals fou Archaeopteryx, el qual tot i no ser un gran volador, ja presentava les plomes com les de les aus modernes. Probablement molts més dinosaures estaven coberts de plomes o protoplomes, però en aquesta entrada només hem mentat les espècies de les que es tenen proves irrefutables de que en presentaven. Com hem vist, el camí fins a arribar a les plomes actuals fou llarg i va donar lloc a una gran diversitat d’espècies de dinosaures, però després de que un meteorit extingís pràcticament la vida a la Terra fa 65 milions d’anys, només un grup de dinosaures emplomallats sobrevisqué i prosperà.
Fòssil de Archaeopteryx lithographica de finals del Juràssic trobat al sud d’Alemanya. Foto de James L. Amos.
T’ha agradat? Recorda valorar la nostra feina votant-nos als Premis Bitácoras com a millor blog d’Educació i Ciència! Moltíssimes gràcies.
Referències
Per a l’elaboració d’aquesta entrada s’han utilitzat les següents referències:
Recently, the BBC documentary series “Planet Dinosaur” has premiered on TVE2. In this series the latest paleontological discoveries concerning the biology of dinosaurs are explained. On my last entry we talked about the theropod dinosaurs, one of which is the spinosaur, one of the largest predators that have ever existed. On this entry I’m going to explain some of the facts that paleontology has revealed about the lifestyle of this creature.
TAXONOMY
The spinosaur (scientific name Spinosaurus aegyptiacus) belonged to the Spinosauridae family, a group of specialized theropods which appeared during the late Jurassic and became extinct about 93 million years ago during the late Cretaceous. This group was characterized by being relatively large theropods, with conic teeth and long snouts similar to crocodiles, and elongated neural spines through its back forming a sail-like structure (that’s where the name “Spinosauridae“ comes from, meaning spine reptiles).
Comparition of the different sizes of various spinosaurids by Scott Hartman. From right to left: Irritator challengeri, Baryonyx walkeri, Suchomimus tenerensis and Spinosaurus aegyptiacus.
Some of the more famous members on this family are, the Baryonyxfrom Europe, which had long curved claws on its hands to capture the fish it fed on, similar to its close relative the Suchomimusfrom northern Africa. Furthermore, there was the smaller Irritatorof about 3 metres tall found in Brasil and finally, the Spinosaurusfrom northern Africa, which measuring between 12 and 18 metres long and wheighing between 7 and 20 tons, was one of the biggest predators to ever walk on land.
HABITAT AND DISTRIBUTION
The genus Spinosaurus was distributed in the zone of what is now the north of Africa. This genus lived during the Cretaceous, appearing about 112 million years ago and disappearing about 97 million years ago.
Map of the World 94 million years ago by Joshua Doubek, during the middle Cretaceous period.
During that period, the northern part of Africa was a very humid zone with high temperatures and lots of wetlands. Spinosaurs probably lived in areas with large rivers and mangrove forests next to the sea, where tidal movements flooded its habitat during certain seasons of the year. This is in accordance with the vision that spinosaurids preferred wet semiaquatic habitats with plenty of great fish to prey upon.
Reconstruction from 2010 of Spinosaurus aegyptiacus by Dmitry Bogdanov.
Currently there are two possible spinosaur species. The most famous is Spinosaurus aegyptiacus from Egypt, the species of which we have more information. A possible second species is Spinosaurus maroccanus from Morocco, which some authors consider simply as a subpopulation of Spinosaurus aegyptiacus.
FUNCTION OF NEURAL SPINES
Spinosaurs were discovered in 1912 from a fossil which included its characteristic dorsal spines. These spines grow up to a length ten times that of the vertebra from which they emerged.
The scarcity of spinosaur fossils means that the function of the spines is still a mystery for science, although there are some hypothesis. One of these is that the spines formed a “sail” along the back of the animal which was highly irrigated and helped the animal’s thermoregulation, as such a big animal probably would have had problems losing heat. Therefore its sail would have helped the spinosaur to evade overheating, orienting it towards fresh winds to cool down.
Reconstruction of the skeleton of a subadult spinosaur (Japan Museum, photo by Kabacchi).
Another hypothesis tells us that the spines held a hump-like structure similar to that of camels, which the animal would have used as a fat reserve system to store fat to withstand periods with little available feeding resources.
Lots of paleontologists think that both hypothesis could be correct and that the spinosaur used the sail both to regulate its body temperature and also to store fat to resist periods of low prey abundance. It is also possible that the sail made the spinosaur appear bigger than it actually was and that they used it during mating rituals similar to those of the modern peacock.
FEEDING STRATEGIES
The Spinosaurus‘s skull shows adaptions to a piscivorous diet. The snout is longer and slender than on other theropods. Aside from this, observing the snout of Spinosaurus it has been seen that it presents a series of little holes similar to those found on crocodiles. It is thought that these structures indicate the presence of pressure receptors which helped them detect the movement of their preys underwater.
Upper jaw of Spinosaurus from the Museo di Storio Naturale di Milano, where the holes which possibly contained the pressure receptors can be seen.
While the teeth of most carnivorous theropods where curved and serrated on their posterior part to tear flesh, spinosaur teeth were conic in shape and had no serration, more similar to those of crocodiles. These teeth were more useful for catching and holding fast and slippery prey and to prevent them from escaping (for example, a fish). Also, various Spinosaurus fossils have been found to have between their theeth scales and bones of large prehistoric fish which probably populated many rivers during the Cretaceous period.
Reconstruction by Joschua Knüppe of two Mawsonia species, the rests of which have been found between the teeth of Spinosaurus.
Nevertheless, it is generally believed that the spinosaur was probably an opportunistic predator, feeding mainly on fish, also hunting small dinosaurs when it had the opportunity and stealing prey from smaller predators using its great size to intimidate them.
POSTURE AND LIFESTYLE
Spinosaurs have traditionally been represented as bipedal animals, as most similarly-sized theropods have. Eventhough most fossils are actually pretty incomplete, it is known that its forelegs were more developed than in most theropods, having long curved claws.
Traditionally it was thought that Spinosaurus hunted in a manner similar to a grey heron, roaming through zones of shallow water, sinking its long snout underwater to detect prey using the pressure receptors, and catching fish with its jaws. It then, probably used its front legs as hands to tear its prey to small pieces easy to swallow.
Reconstruction by Joschua Knüppe of Spinosaurus aegyptiacus in hunting posture.
At the end of 2014 a new Spinosaurus fossil was discovered which has changed the view we had on this animal. For the first time, scientists found a fossil which shows the structure of the hind legs of this dinosaur and they have observed a number of characteristics not found in any other theropod not even in other spinosaurids. This fossil shows that the hind legs of Spinosaurus were much more massive than those of other theropod dinosaurs, in which the bones are usually hollow to make them more agile (like present day birds). Also, in this fossil the hind legs are actually much shorter in relation to the size of the animal than in any other theropod, leading some scientists to think that Spinosaurus was actually a quadrupedal animal. This has made some paleontologist think that maybe the lifestyle of the spinosaurs was much more similar to that of a crocodile and that they spent much more time living in water than on land, making the Spinosaurus the first known aquatic dinosaur.
Reconstruction by Rodrigo Vega of Spinosaurus based on the skeleton found in 2014.
Anyway, many paleontologists argue that the biology of a species cannot be based on a single fossil and advise caution when generalizing to the whole species (the fossil could belong to an adult and a juvenile that died together or could even come from an individual which had suffered some kind of embryonic malformation that kept its legs from developing normally). Paleontology is a science in which with every new discovery we can unravel the tree of life and the evolution of the different groups of living beings. With a little of luck, future discoveries will enable us to clarify the anatomy of Spinosaurus aegyptiacus and define the lifestyle of such a unique and extraordinary reptile.
REFERENCES
The following sources have been consulted in the elaboration of this entry:
Ifyou enjoyed thisarticle, pleaseshare iton social networks to spread it. The aim of theblog, after all, is to spread scienceand reach as many people aspossible.
This publication is under a Creative Commons License:
Recentment s’ha estrenat a La 2 de TVE la sèrie documental de la BBC “Planeta Dinosaure” on s’expliquen els descobriments paleontològics més recents relacionats amb la biologia dels dinosaures. En la entrada anterior vam parlar dels dinosaures teròpodes entre els qual es troba l’espinosaure, un dels depredadors més grans que ha existit mai. En aquesta entrada explicaré algunes de les coses que la paleontologia ha revelat sobre l’estil de vida d’aquest animal.
TAXONOMIA
L’espinosaure (de nom científic Spinosaurus aegyptiacus) pertanyia a la família Spinosauridae, un grup de teròpodes especialitzats que va aparèixer a finals del Juràssic i desaparegué fa uns 93 milions d’anys a finals del Cretàcic. Aquest grup es caracteritzava per ser teròpodes relativament grans, amb dents còniques i morros allargats semblants als dels cocodrils i espines neurals allargades recorrent l’esquena formant una estructura en forma de “vela” (d’aquí ve el nom “Spinosauridae”, rèptils amb espines).
Comparació de la mida de diferents espinosàurids per Scott Hartman. De dreta a esquerra: Irritator challengeri, Baryonyx walkeri, Suchomimus tenerensis i Spinosaurus aegyptiacus.
Els membres més coneguts d’aquesta família són el Baryonyxd’Europa, que tenia unes grans urpes corbades a les potes del davant per a capturar els peixos dels quals s’alimentava de forma semblant al seu parent proper, el Suchomimusdel nord d’Àfrica. A més, tenim al petit Irritatord’uns 3 metres d’alt descobert al Brasil i finalment, a l’Spinosaurusdel nord d’Àfrica, que mesurant entre 12 i 18 metres de llarg i pesant entre 7 i 20 tones, va ser un dels depredadors més grans que ha caminat mai per terra.
HÀBITAT I DISTRIBUCIÓ
El gènere Spinosaurus es trobava distribuït per la zona corresponent al que actualment és el nord d’Àfrica. Aquest gènere va viure durant el Cretàcic, apareixent fa uns 112 milions d’anys i extingint-se fa uns 97 milions d’anys.
Mapa del món fet per Joshua Doubek de fa 94 milions d'anys, a mitjans del Cretàcic.
Durant aquest període, el nord d’Àfrica era una zona humida d’altes temperatures i zones pantanoses. Segurament els espinosaures vivien en àrees amb grans rius i manglars a prop de la costa, on les marees inundaven el seu hàbitat en diferents èpoques de l’any. Això concorda amb la visió de que els espinosàurids preferien zones humides amb abundància de grans peixos dels que alimentar-se.
Reconstrucció del 2010 de Spinosaurus aegyptiacus per Dmitry Bogdanov.
Actualment hi ha dues possibles espècies d’espinosaures. La més famosa és Spinosaurus aegyptiacus de Egipte, que és l’espècie de la qual més coses sabem. Una segona espècie possible és Spinosaurus maroccanus del Marroc, que alguns consideren simplement una subpoblació de Spinosaurus aegyptiacus.
FUNCIÓ DE LES ESPINES DORSALS
Els espinosaures van ser descoberts el 1912 per un fòssil que incloïa les característiques espines dorsals. Aquestes espines arribaven a fer 10 vegades la longitud de les vèrtebres de les quals emergien.
La escassetat de fòssils d’espinosaures ha provocat que la funció de les espines sigui encara un misteri per la ciència. Tot i així, existeixen vàries hipòtesis. Una d’aquestes és que les espines formaven una “vela” al llarg de l’esquena altament irrigada amb vasos sanguinis que ajudava a l’animal en la termoregulació, ja que probablement un animal tant gran tingués problemes per a perdre calor. La vela hauria ajudat a l’animal a evitar el sobreescalfament orientat-la cap a la direcció del vent per a refrescar-se.
Reconstrucció d'esquelet d'espinosaure subadult (Museu de Japó, foto de Kabacchi).
Una altra hipòtesi diu que les espines formaven una estructura més semblant a la gepa d’un camell que l’animal utilitzava per a emmagatzemar greix per a suportar períodes d’escassetat de recursos.
Molts paleontòlegs creuen que ambdues hipòtesis podrien ser correctes i que l’espinosaure utilitzés la vela tant per a regular la seva temperatura corporal i a més que també hi emmagatzemés greix per a resistir períodes de falta d’aliment. A més, la vela segurament feia que l’espinosaure semblés encara més gran del que era i també que la fessin servir en els rituals d’aparellament de forma semblant als paons actuals.
ALIMENTACIÓ
El crani de Spinosaurus mostra adaptacions a una dieta piscívora. El musell és més allargat i prim que en altres teròpodes. Apart d’això, en observar el morro del Spinosaurus s’ha observat que presenten un seguit de petits forats semblants als que presenten els cocodrils. Es creu que aquestes estructures indiquen la presència de receptors de pressió que els permetia detectar el moviment de les seves preses a sota de l’aigua.
Mandíbula superior de Spinosaurus del Museo di Storio Naturale di Milano, on s'observen els forats que possiblement contenien els receptors de pressió.
Mentre que les dents de la majoria de teròpodes carnívors eren corbades i presentaven una serra a la part posterior per a tallar la carn, les dents de l’espinosaure eren còniques i sense serra, més semblants a les d’un cocodril. Aquestes dents eren més útils per atrapar i impedir que preses ràpides i relliscoses se’ls escapessin (com per exemple un peix). A més, en varis fòssils de Spinosaurus s’han trobat entre les seves dents restes d’escates i ossos de grans peixos prehistòrics que segurament poblaven els rius del Cretàcic.
Reconstrucció feta per Joschua Knüppe de dues espècies de Mawsonia, uns peixos dels quals s'han trobat restes entre les dents de Spinosaurus.
Tanmateix es creu que l’espinosaure devia ser un depredador oportunista, alimentant-se principalment de peixos, però caçant petits dinosaures quan podia o robant les preses a altres depredadors aprofitant-se de la seva mida.
POSTURA I ESTIL DE VIDA
Els espinosaures tradicionalment se’ls ha representat com a animals bípedes, igual que la majoria de teròpodes de mida semblant. Tot i que la majoria de fòssils trobats són bastant incomplets, es sap que les potes del davant estaven més desenvolupades que les de la majoria de teròpodes i que presentaven unes urpes llargues i corbades.
Tradicionalment es creia que Spinosaurus caçava d’una forma semblant al bernat pescaire, rondant per zones d’aigües poc profundes, enfonsant el seu llarg morro per a detectar a les preses mitjançant els receptors de pressió i capturant-la amb les mandíbules. Segurament, també utilitzava les potes del davant per a acabar de desmembrar les preses en troços més petits i menjar-se-les.
Reconstrucció feta per Joschua Knüppe de Spinosaurus aegyptiacus en postura de caça.
A finals del 2014 es va descobrir un nou fòssil de Spinosaurus que ha canviat la visió que es tenia sobre aquest animal. Per primer cop s’ha trobat un fòssil que mostra les potes del darrera d’aquest dinosaure i s’han vist un seguit de característiques que no es troben en cap altre teròpode i ni tan sols en cap altre espinosàurid. Aquest fòssil ens mostra que les potes del darrere eren molt més massisses que en altres dinosaures teròpodes, els quals solen tenir els ossos buits fent-los més lleugers (com en els ocells actuals). A més, en aquest fòssil les potes del darrere són molt més curtes en relació a la mida de l’animal que en cap altre teròpode, cosa que ens porta a pensar que Spinosaurus probablement fós un animal quadrúpede. Això ha provocat que alguns científics creguin que l’estil de vida d’aquests dinosaures era més semblant al del cocodril i que aquests passarien molt més temps vivint a l’aigua que a terra, convertint a l’espinosaure en el primer dinosaure aquàtic conegut.
Reconstrucció feta per Rodrigo Vega de Spinosaurus basant-se en el esquelet descobert el 2014.
De tota manera, molts paleontòlegs argumenten que no es pot basar la biologia d’una espècie en un únic fòssil i aconsellen precaució a l’hora de fer generalitzacions a tota l’espècie (el fòssil podria tractar-se d’una barreja d’ossos d’individus adults i juvenils o fins i tot podria ser un individu que va patir una mutació congènita que fes que les potes posteriors no es desenvolupessin correctament). La paleontologia és una ciència que amb cada nou descobriment va desentranyant l’arbre de la vida i la evolució dels diferents grups d’éssers vius. Amb una mica de sort, descobriments posteriors ens permetran aclarir l’anatomia del Spinosaurus aegyptiacus per així poder acabar definint l’estil de vida d’aquest rèptil tant únic i extraordinari.
REFERÈNCIES
S’han consultat les següents fonts per a elaborar els continguts d’aquesta entrada:
Si t’ha agradat aquest article, si us plau comparteix-lo a les xarxes socials per a fer-ne difusió, doncs l’objectiu del blog, al cap i a la fi, és divulgar la ciència i que arribi al màxim de gent possible.
Aquesta publicació té una llicència Creative Commons:
Recientemente se ha estrenado en La 2 de TVE la serie documental de la BBC “Planeta Dinosaurio”, en la que se explican los descubrimientos paleontológicos más recientes relacionados con la biología de los dinosaurios. En la entrada anterior hablamos sobre los dinosaurios terópodos entre los cuales se encuentra el espinosaurio, uno de los mayores depredadores que han existido. En ésta entrada explicaré algunas de las cosas que la paleontología ha revelado sobre el estilo de vida de este animal.
TAXONOMÍA
El espinosaurio (de nombre científico Spinosaurus aegyptiacus) pertenecía a la familia Spinosauridae, un grupo de terópodos especializados que apareció a finales del Jurásico y desapareció hace unos 93 miliones de años a finales del Cretácico. Este grupo se caracterizaba por ser terópodos relativamente grandes, con dientes cónicos y hocicos alargados parecidos a los de los cocodrilos y espinas neurales alargadas recorriendo la espalda formando una estructura en forma de “vela” (de aquí viene el nombre “Spinosauridae”, reptiles con espinas).
Comparación del tamaño de diferentes espinosáuridos por Scott Hartman. De derecha a izquierda: Irritator challengeri, Baryonyx walkeri, Suchomimus tenerensis y Spinosaurus aegyptiacus.
Los miembros más conocidos de esta familia son el Baryonyxde Europa, que tenía unas grandes garras curvadas en las patas delanteras para capturar los peces de los que se alimentaba de forma similar a su primo cercano, el Suchomimusdel norte de África. Además, tenemos al pequeño Irritatorde unos 3 metros de alto descubierto en Brasil y finalmente, al Spinosaurusdel norte de África, que midiendo entre 12 y 18 metros de largo y pesando entre 7 y 20 toneladas, fue uno de los depredadores más grandes que ha caminado nunca por tierra.
HÁBITAT Y DISTRIBUCIÓN
El género Spinosaurus se encontraba distribuido por la zona correspondiente a lo que actualmente es el norte de África. Este género vivió durante el Cretácico, apareciendo hace unos 112 millones de años y extinguiéndose hace unos 97 millones de años.
Mapa del mundo de hace 94 millones de años, a mediados del Cretácico, hecho por Joshua Doubek.
Durante este periodo, el norte de África era una zona húmeda, de altas temperaturas y zonas pantanosas. Seguramente los espinosaurios vivían en áreas con grandes ríos y manglares, cercanas al mar, donde las mareas inundaban su hábitat en diferentes épocas del año. Esto concuerda con la visión que se tiene de que los espinosáuridos preferían zonas húmedas con abundancia de grandes peces de los que alimentarse.
Reconstrucción del 2010 de Spinosaurus aegyptiacus por Dmitry Bogdanov.
Actualmente hay dos posibles especies de espinosaurios. La más famosa es Spinosaurus aegyptiacus de Egipto, que es la especie de la cual más cosas sabemos. Una segunda especie posible es Spinosaurus maroccanus de Marruecos, a la que algunos consideran simplemente como una subpoblación de Spinosaurus aegyptiacus.
FUNCIÓN DE LAS ESPINAS DORSALES
Los espinosaurios fueron descubiertos en 1912 a partir de un fósil que incluía las características espinas dorsales. Estas espinas llegaban a medir 10 veces la longitud de las vértebras de las cuáles emergían.
La escasez de fósiles de espinosaurio ha provocado que la función de las espinas sea aún un misterio para la ciencia. Aun así, existen varias hipótesis al respecto. Una de éstas es que las espinas formaban una “vela” a lo largo del dorso altamente irrigada con vasos sanguíneos que ayudarían al animal en la termorregulación, ya que probablemente un animal de este tamaño tendría problemas para perder calor. Por lo tanto, la vela habría ayudado al animal a evitar el sobrecalentamiento orientándola hacia la dirección del viento para refrescarse.
Reconstrucción de esqueleto de espinosaurio subadulto (Museo de Japón, foto de Kabacchi).
Otra hipótesis dice que las espinas formaban una estructura parecida a la joroba de un camello, que el animal utilizaba para almacenar grasa para soportar períodos de escasez de recursos.
Muchos paleontólogos creen que ambas hipótesis podrían ser correctas y que el espinosaurio utilizaría la vela tanto para regular su temperatura corporal como para almacenar grasa para resistir períodos de falta de alimento. Además, la vela seguramente hacía que el espinosaurio pareciese aún más grande de lo que era y que también la utilizase en los rituales de apareamiento de forma parecida a los pavos reales actuales.
ALIMENTACIÓN
El cráneo de Spinosaurus muestra adaptaciones a una dieta piscívora. El hocico es más alargado y delgado que en otros terópodos. Aparte de esto, al observar el hocico del Spinosaurus se ha observado que presenta un seguido de pequeños agujeros parecidos a los que presentan los cocodrilos. Se cree que estas estructuras indican la presencia de receptores de presión que les permitía detectar el movimiento de sus presas bajo el agua.
Mandíbula superior de Spinosaurus del Museo di Storio Naturale di Milano, en la que se observan los agujeros que posiblemente contenían los receptores de presión.
Mientras que los dientes de la mayoría de terópodos carnívoros eran curvados y presentaban un borde posterior aserrado para cortar la carne, los dientes del espinosaurio eran cónicos y sin sierra, más parecidos a los de un cocodrilo. Estos dientes eran más útiles para atrapar e impedir que presas rápidas y resbaladizas se les escaparan (cómo por ejemplo un pez). Además, en varios fósiles de Spinosaurus se han encontrado entre sus dientes restos de escamas y huesos de grandes peces prehistóricos que seguramente poblaban los ríos del Cretáceo.
Reconstrucción hecha por Joschua Knüppe de dos especies de Mawsonia, unos peces de los cuales se han encontrado restos entre los dientes de Spinosaurus.
Aun así, se cree que el espinosaurio debía ser un depredador oportunista, alimentándose principalmente de peces, pero cazando pequeños dinosaurios cuando podía o robando las presas a otros depredadores aprovechándose de su tamaño.
POSTURA Y ESTILO DE VIDA
Tradicionalmente se han representado a los espinosaurios como animales bípedos, igual que a la mayoría de terópodos de similar tamaño. Aunque la mayoría de fósiles encontrados son bastante incompletos, se sabe que las patas delanteras estaban más desarrolladas que las de la mayoría de terópodos y que presentaban unas garras largas y curvadas.
Tradicionalmente se creía que el Spinosaurus cazaba de forma similar a una garza real, rondando por zonas de aguas poco profundas, hundiendo su largo hocico para detectar a las presas mediante los receptores de presión y capturándola con las mandíbulas. Seguramente, también utilizaba las patas de delante para acabar de desmembrar a las presas en trozos más pequeños y comérselas.
Reconstrucción hecha por Joschua Knüppe de Spinosaurus aegyptiacus en postura de caza.
A finales del 2014 se descubrió un nuevo fósil de Spinosaurus que ha cambiado la visión que se tenía sobre este animal. Por primera vez se ha encontrado un fósil que muestra las patas traseras de este dinosaurio y se ha visto un conjunto de características que no se encuentran en ningún otro terópodo y ni siquiera en ningún otro espinosáurido. Este fósil nos muestra que las patas de atrás eran mucho más macizas que en otros dinosaurios terópodos, los cuales suelen tener los huesos huecos haciéndolos más ligeros (como en las aves actuales). Además, en este fósil las patas traseras son mucho más cortas en relación al tamaño del animal que en ningún otro terópodo, cosa que lleva a pensar que Spinosaurus probablemente fuese un animal cuadrúpedo. Esto ha provocado que algunos científicos crean que el estilo de vida de estos dinosaurios era más parecido al de un cocodrilo y que estos pasarían mucho más tiempo viviendo en el agua que en tierra, convirtiendo al espinosaurio en el primer dinosaurio acuático conocido.
Reconstrucción hecha por Rodrigo Vega de Spinosaurus basándose en el esqueleto descubierto en 2014.
Aun así, muchos paleontólogos argumentan que no se puede basar la biología de un animal en un único fósil y aconseja precaución a la hora de hacer generalizaciones a toda la especie (el fósil podría tratarse de una mezcla de huesos de individuos adultos y juveniles o incluso podría ser un individuo que sufriese una mutación congénita que hiciese que las patas posteriores no se desarrollaran correctamente). La paleontología es una ciencia en la que con cada nuevo descubrimiento va desentrañando el árbol de la vida y la evolución de los distintos grupos de seres vivos. Con un poco de suerte, descubrimientos posteriores nos permitirán esclarecer la anatomía del Spinosaurus aegyptiacus para así poder acabar definiendo el estilo de vida de este reptil tan único y extraordinario.
REFERENCIAS
Se han consultado las siguientes fuentes en la elaboración de los contenidos de esta entrada:
Si te ha gustado este artículo, por favor compártelo en las redes sociales. El objetivo del blog, al fin y al cabo, es divulgar la ciencia y llegar a tanta gente como sea posible. Siéntete libre de dar tus comentarios.
Esta publicación tiene una licencia Creative Commons:
Los dinosaurios (superorden Dinosauria, que significa “reptiles temibles”) son un grupo de reptiles que dominaron todos los ecosistemas terrestres durante el Mesozoico (Era Secundaria o “la Era de los Reptiles”). Aun así, mucha gente todavía no tiene claro qué es un dinosaurio y qué no lo es, y generaliza el término “dinosaurio” para referirse a todos los reptiles que evolucionaron durante la Era Secundaria. En esta entrada intentaré hacer un pequeño repaso de los diferentes grupos de reptiles que aparecieron durante el Mesozoico y explicaré por encima, la clasificación de los diferentes grupos de dinosaurios y algunas de sus adaptaciones.
REPTILES DEL MESOZOICO: DINOSAURIOS Y MUCHOS MÁS
Los dinosaurios aparecieron después de un fenómeno de extinción masiva que ocurrió hace unos 251 millones de años (extinción del Pérmico-Triásico). Éste fenómeno eliminó el 96% de las especies marinas y el 70% de las especies terrestres del momento, dejando muchos nichos ecológicos vacios que podrían ser ocupados por nuevas especies.
Gráfico modificado de Rohde & Muller (2005) mostrando las grandes extinciones masivas. La zona más oscura indica el período correspondiente al Mesozoico.
Durante el Triásico (a principios del Mesozoico) evolucionaron muchos grupos de reptiles diferentes. Entre estos grupos encontramos a los dinosaurios, los cuales en esos momentos estaban lejos de ser los vertebrados terrestres dominantes. Entre algunos de los otros grupos de reptiles no dinosaurios de esa época encontramos: los rauisúquidos (clado Rauisuchia) terrestres y grupos totalmente marinos como los sauropterigios (superorden Sauropterygia) y los ictiopterigios (superorden Ichthyopterygia).
Reconstrucciones de Dmitry Bogdanov de Prestosuchus (un rauisúquido, arriba), de Nichollsia (un sauropterigio, abajo izquierda) y de Platypterigius (un ictiopterigio, abajo derecha).
Una segunda extinción masiva a finales del Triásico y a principios del Jurásico acabó con muchos de los reptiles dominantes, lo que permitió la expansión y evolución de los hasta ahora pequeños dinosaurios y de nuevos grupos como los crocodilomorfos (superorden Crocodylomorpha, antepasados de los cocodrilianos), los pterosaurios voladores (orden Pterosauria).
Reconstrucciones de Dmitry Bogdanov de Dakosaurus (un crocodilomorfo, arriba) y de Scaphognathus (un pterosaurio, abajo).
Como podemos ver, los dinosaurios sólo son uno de los muchos grupos de reptiles que evolucionaron durante el Mesozoico. Durante el Jurásico, los dinosaurios se diversificaron mucho, aunque prácticamente todas las especies eran exclusivamente terrestres. Éstos, dominaron los ecosistemas terrestres de la Tierra hasta su práctica extinción hace 65 millones de años a finales del Cretácico.
CLASIFICACIÓN DE LOS DINOSAURIOS
Los primeros dinosaurios evolucionaron hace unos 231 millones de años a mediados del Triásico. Éstos, eran animales pequeños y se caracterizaban por la postura de sus patas que, a diferencia del resto de los reptiles, eran verticales y elevaban su cuerpo del suelo. Esto les permitía una mayor movilidad y un estilo de vida más activo.
Arriba: Esqueleto de Eoraptor, uno de los dinosaurios más antiguos que se conocen (Museo de Japón, foto de Kentaro Ohno). Abajo: Esquemas de la postura típica de la mayoría de reptiles (izquierda) y la característica de los dinosaurios (derecha).
Posteriormente, los dinosaurios se separaron en dos órdenes diferentes: los Saurischiay los Ornithischia. Estos dos grupos se diferenciaban en la estructura de su pelvis; los saurisquios conservaron una pelvis parecida a la del resto de reptiles, mientras que los ornitisquios evolucionaron una pelvis superficialmente más parecida a la de los pájaros.
Esquema mostrando la estructura de la pelvis de los Saurischia (izquierda) y de los Ornithischia (derecha). Representa que los animales están mirando a la izquierda.
ORNITISCHIA: PELVIS DE AVE
Árbol evolutivo de los Ornithischia, modificado de Zureks.
Ornithopoda (“pies de ave”): Los ornitópodos fueron el grupo de ornitisquios más diverso, que se caracterizaban por tener pies con tres dedos parecidos a los de los pájaros. Eran herbívoros que podían combinar la marcha bípeda y la cuadrúpeda. Entre ellos se encuentra el Iguanodon, uno de los primeros dinosaurios descubiertos por la ciencia.
Pie de Iguanodon (derecha) y reconstrucción de O. C. Marsh (1896).
Los ornitópodos adquirieron muchas adaptaciones diferentes; algunos grupos tenían picos de pato para alimentarse de vegetación acuática, otros desarrollaron manos muy especializadas con un pulgar en forma de puñal para defenderse y un meñique oponible para asir la vegetación de la que se alimentaban. Muchos grupos desarrollaron crestas óseas que se cree que les servían tanto para diferenciarse entre miembros de distintas especies, como para comunicarse entre ellos.
Reconstrucción de Parasaurolophus (autor desconocido), un ornitópodo que presentaba una gran cresta hueca que le servía para amplificar los sonidos que producía.
Marginocephalia (“cabeza flanqueada”): Los llamados marginocéfalos fueron un grupo de dinosaurios herbívoros emparentados con los ornitópodos caracterizados por una amplia osificación del cráneo. Éstos se pueden dividir en dos grupos:
Los paquicefalosaurios (suborden Pachycephalosauria, “reptiles de cabeza gruesa”) eran bípedos que presentaban el cráneo muy grueso y flanqueado por una serie de osteodermos (osificaciones recubiertas de queratina). Se cree que los paquicefalosaurios resolvían las disputas territoriales y resolvían los conflictos reproductores chocando las cabezas de forma similar a las cabras.
Reconstrucción de Pachycephalosaurus por Jordan Mallon.
Los otros miembros de los marginocéfalos son los ceratópsidos (suborden Ceratopsia, “cara con cuernos”), cuadrúpedos presentaban; un collar óseo que hacía que el cráneo pareciese mucho más grande y el “hueso rostral” que formaba una estructura en forma de pico en el hocico. Muchas especies desarrollaron además, cuernos faciales que podían salir tanto del pómulo, como de encima de los ojos, como del collar óseo.
Reconstrucción de Rubeosaurus por Lukas Panzarin (izquierda) y cráneo de Triceratops (derecha), foto de Zachi Evenor.
Thyreophora (“portadores de escudos”): Este grupo basal de ornitisquios estaba exclusivamente formado por herbívoros cuadrúpedos que presentaban grandes osteodermos que constituían su principal método de defensa. Los tireóforos se pueden dividir en:
Los estegosaurios (suborden Stegosauria, “reptiles con tejas”) fueron grandes dinosaurios herbívoros caracterizados por dos filas de osteodermos que les recorría el dorso desde el cuello hasta la cola y que les servía tanto para su protección cómo para ayudar a la termorregulación. Algunas especies también desarrollaron espinas caudales llamadas “thagomizers” que utilizaban como armas para protegerse de los depredadores.
“Thagomizer” montado en el Denver Museum of Nature and Science (izquierda) y reconstrucción de Stegosaurus de Nobu Tamura (derecha).
Los anquilosaurios (suborden Ankylosauria, “reptiles fundidos”) desarrollaron grandes armaduras óseas que les cubrían todo el cuerpo. Algunos como Ankylosaurus, desarrollaron mazas óseas en el extremo de la cola para defenderse de los depredadores.
Árbol evolutivo de los Saurischia, modificado de Zureks.
Sauropodomorpha (“pies con forma de reptil”): Los sauropodomorfos son más conocidos como “dinosaurios cuellilargos”. Esto se debe a que evolucionaron para alimentarse de los estratos de vegetación más elevados.
Reconstrucción de diferentes sauropodomorfos (de izquierda a derecha): Camarasaurus, Brachiosaurus, Giraffatitan y Euhelopus.
La mayoría de especies se convirtieron en enormes cuadrúpedos con las patas en forma de columna parecidas a las de los elefantes y cuellos largos para llegar a las hojas de los árboles más altos. Las especies más evolucionadas adquirieron tamaños gigantescos como el Amphicoelias, que podía llegar a crecer hasta los 60 metros de longitud.
Theropoda (“pies de bestia”): Este último grupo es conocido por dos motivos. El primero es que este grupo incluye a algunas de las familias más famosas de grandes depredadores, como los Tyrannosauridae y los Dromeosauridae. El segundo motivo es que son el único grupo de dinosaurios con representantes vivos, ya que las aves se incluyen dentro del suborden Theropoda.
Esqueleto de Allosaurus de colecciones de museos americanos (1915).
Todos los terópodos son bípedos y la mayoría de especies mesozoicas eran carnívoras, con dientes renovables y afilados, adaptados a la depredación. Los terópodos presentan una pelvis saurisquia, pero posteriormente los pájaros desarrollaron una pelvis más parecida a la de los dinosaurios ornitisquios.
Reconstrucción de Davide Bonnadonna de los diferentes clados que condujeron a la aparición de los pájaros (de izquierda a derecha): Neotheropoda, Tetanurae, Coelurosauria, Paraves y finalmente el Archaeopteryx, que se cree que fué la primera especie de ave que existió.
Algunas especies presentaban plumas que les ayudaban a la termorregulación. A partir de estos grupos evolucionaron los pájaros a finales del Jurásico.
REFERENCIAS
Se han consultado las siguientes fuentes para elaborar los contenidos de esta entrada:
Si te ha gustadoeste artículo, por favor compárteloen las redes socialesparahacer difusión, puesel objetivo delblog, al fin y al cabo,es divulgarla cienciayque llegueal máximode gente posible.

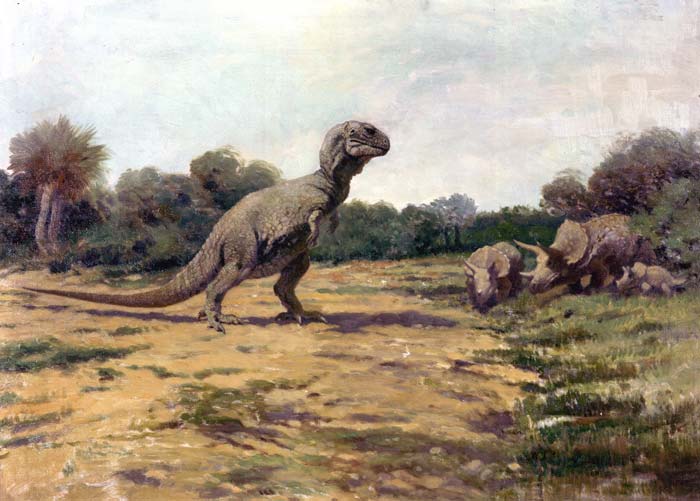
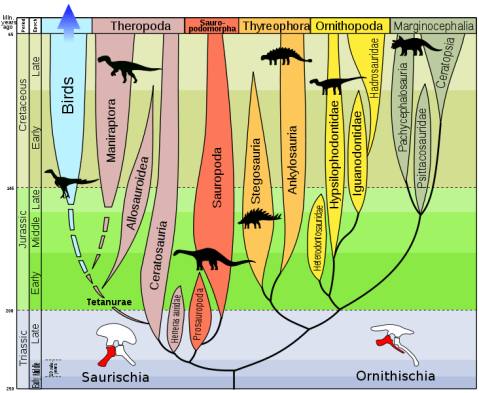


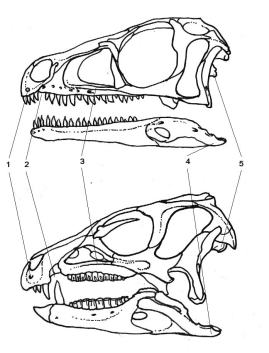

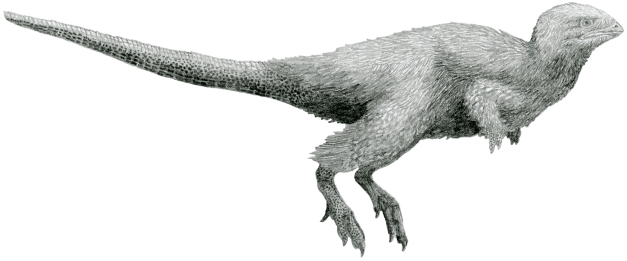
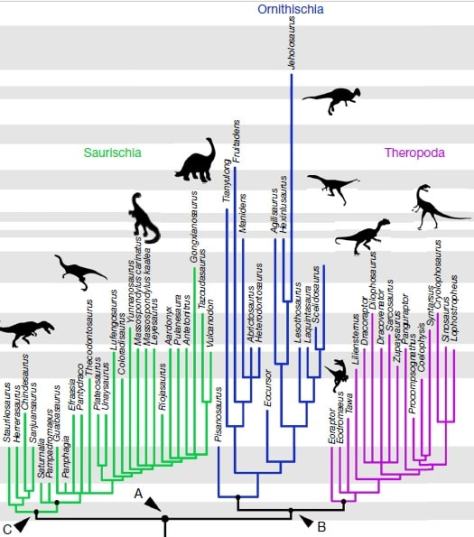
































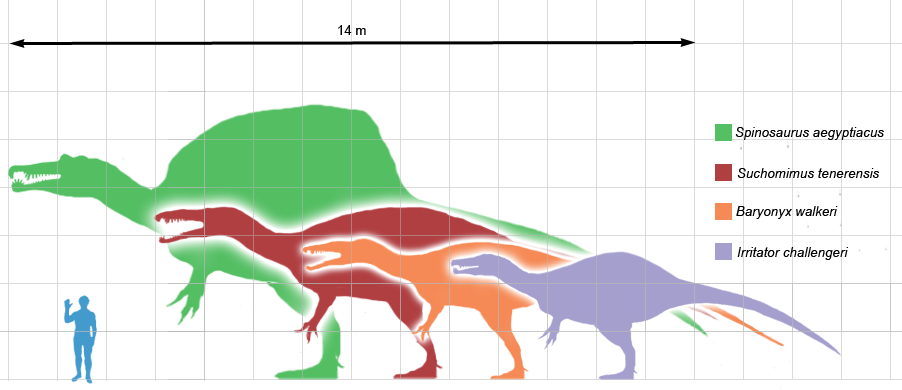
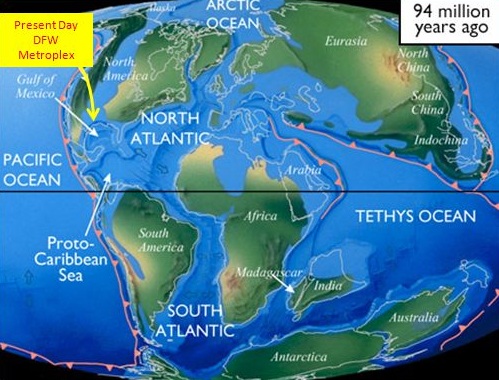
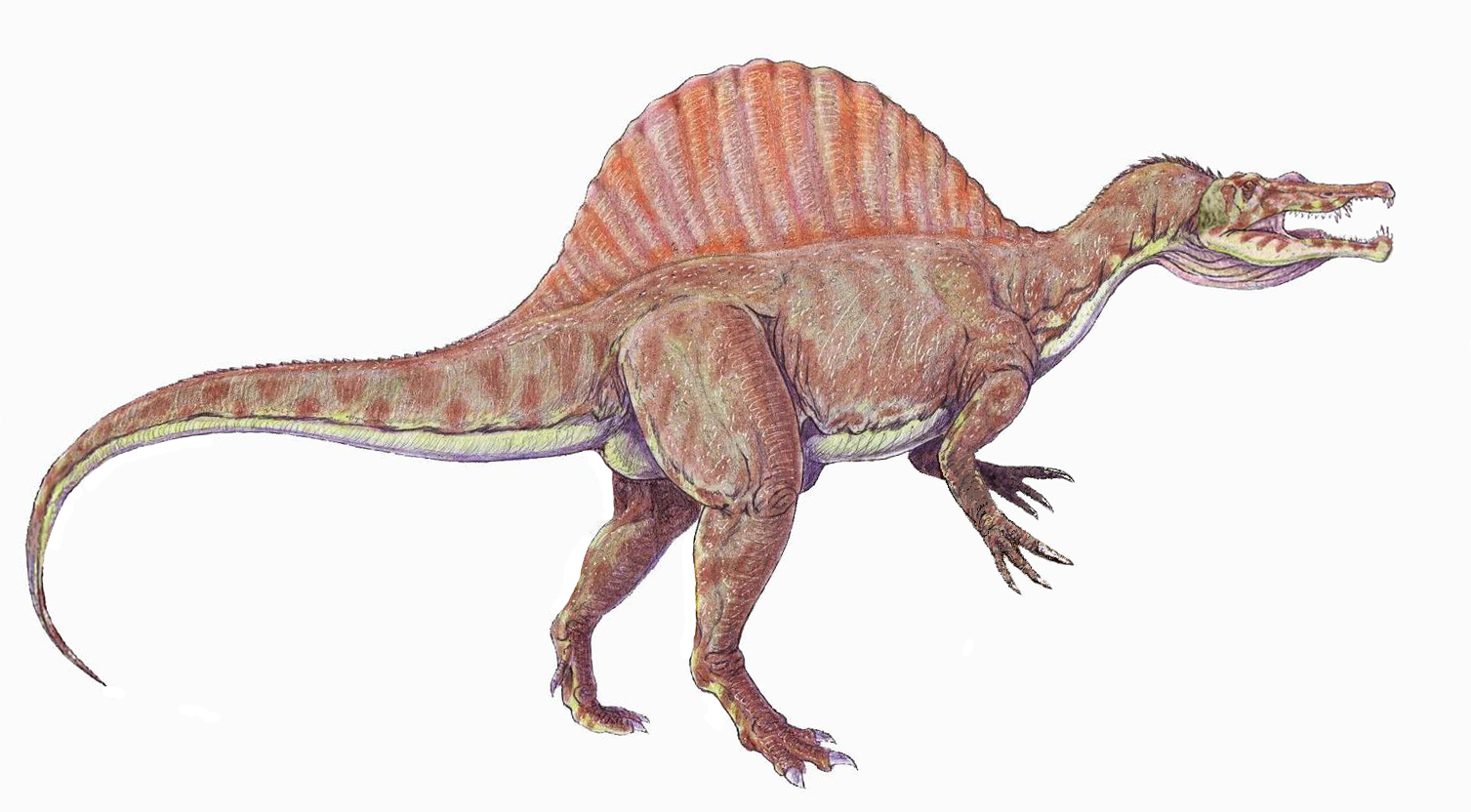



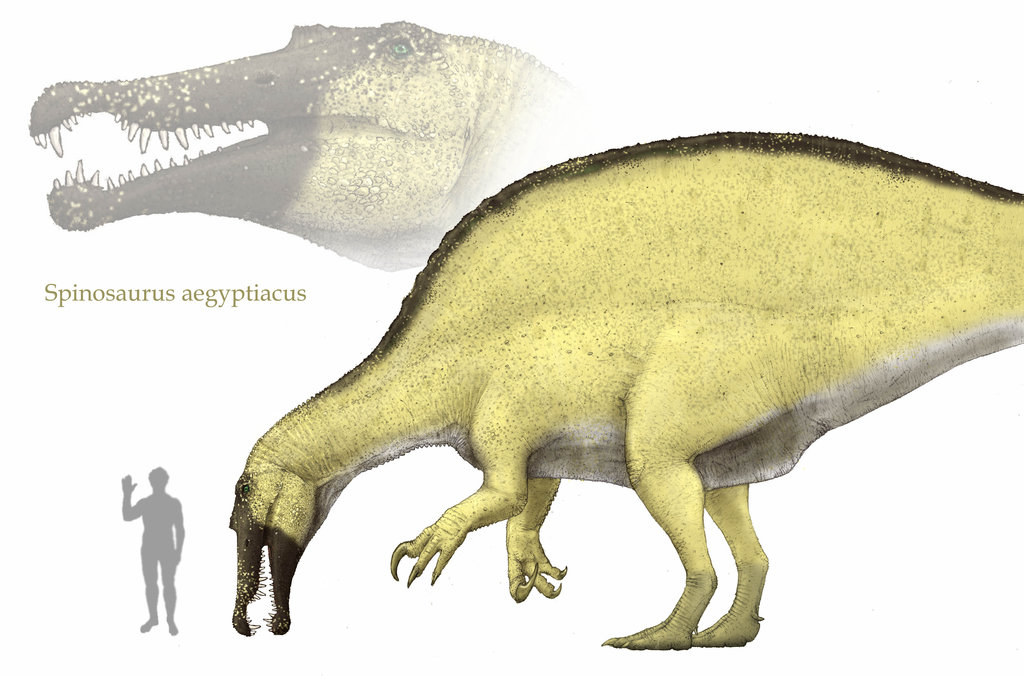




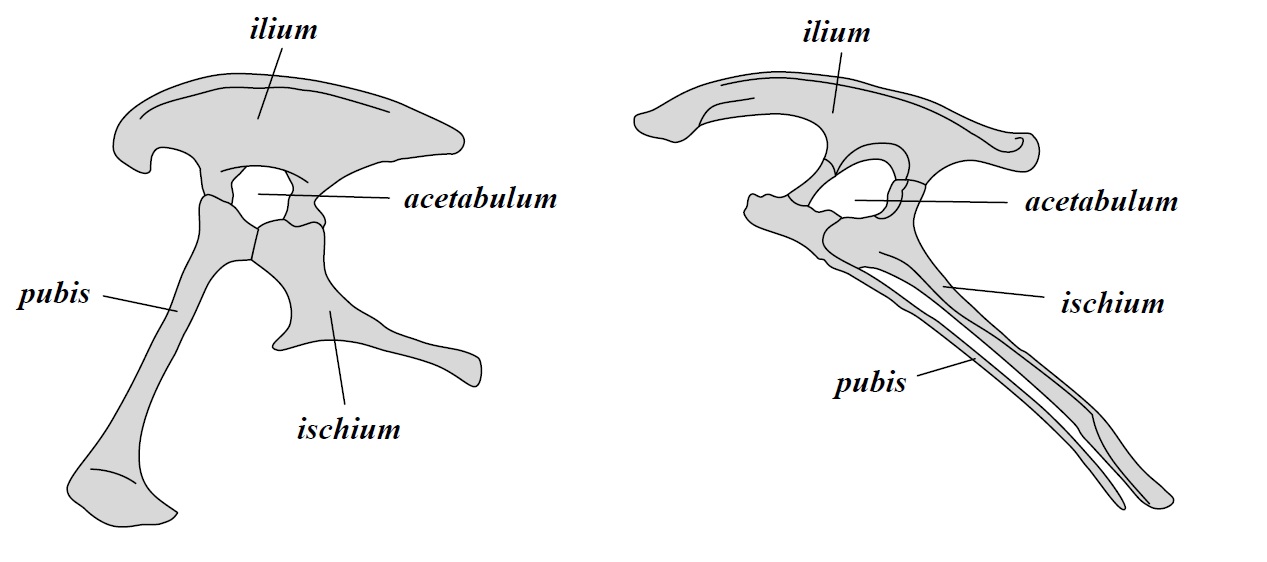
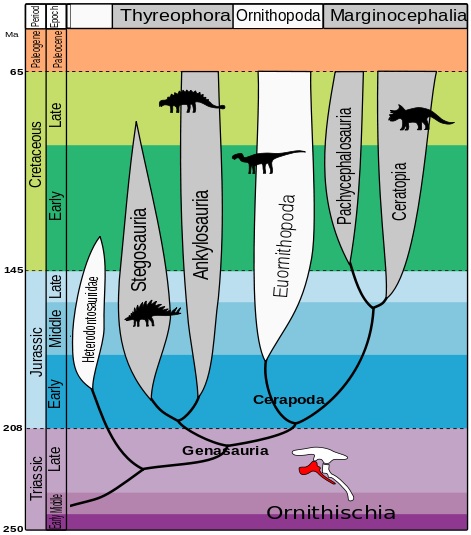
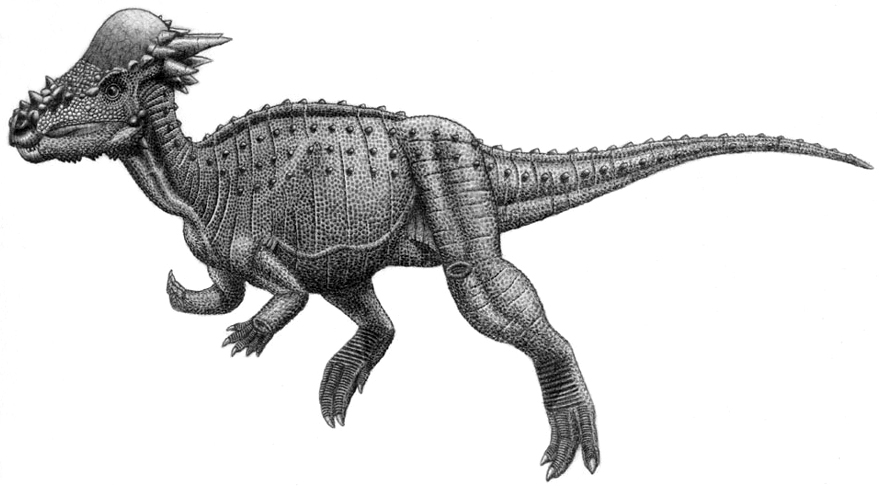



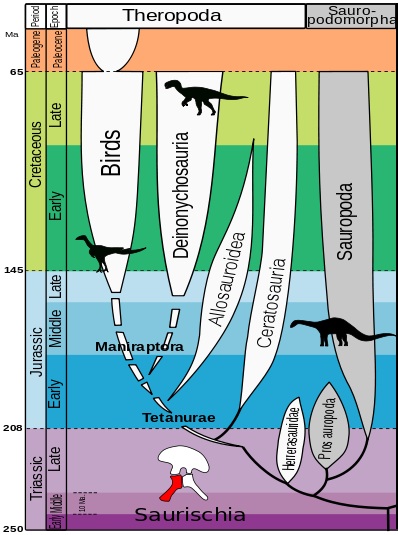


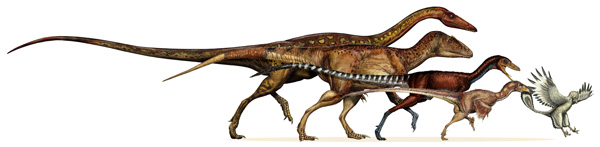


{kind=link}
{kind=link}