

Com s’ho fan alguns insectes, aranyes o llangardaixos per caminar sobre parets llises o de cap per avall i no caure? ¿Per què, si fos real, l’Spiderman no podria enganxar-se a les parets com ho fan aquests animals?
Científics de diferents àrees encara busquen comprendre els mecanismes que fan servir alguns animals per caminar sobre aquest tipus de superfícies sense relliscar o precipitar-se. A continuació, t’expliquem què sap la comunitat científica sobre aquest fenomen.
Animals que caminen per la paret: un repte a la gravetat
La competència per l’espai i els recursos (nínxol ecològic) ha donat lloc a nombroses i increïbles adaptacions al llarg de l’evolució, com la miniaturització.
Quan una superfície és massa llisa, de manera que les ungles, les urpes o les forces de fricció resulten insuficients per a desplaçar-se sobre ella sense caure, entren en joc mecanismes d’adhesió dinàmica: aquells que permeten a l’animal desplaçar-se sobre superfícies verticals llises o de cap per avall enganxant-se i desenganxant-se ràpidament. L’aparició d’estructures adhesives dinàmiques ha permès a diversos animals explotar nous ambients, podent desplaçar-se per caçar o romandre immòbils el temps necessari per fugir dels seus depredadors allà on la majoria tan sols podria estar estable uns pocs segons.

El desenvolupament d’estructures adhesives dinàmiques en les extremitats és típic d’insectes i d’aranyes, d’alguns rèptils com els geckos i certes sargantanes, i d’amfibis com les granotes arborícoles. Puntualment, també s’ha observat en petits mamífers com ratpenats i pòssums, uns marsupials arborícoles procedents d’Austràlia i de certes regions del sud-est asiàtic.
El fet que grups tan diferents d’animals presentin una adaptació similar s’explica per un procés de convergència evolutiva: davant un mateix problema (competència per l’espai i els recursos, elevada pressió de depredació, etc.), l’evolució tendeix a solucions iguals o similars (estructures adhesives per accedir a altres espais).
Els límits de l’adaptació (o per què l’Spiderman no podria caminar per les parets)
Estudiar el mecanisme mitjançant el qual alguns animals caminen sobre superfícies verticals llises o invertides és clau per al desenvolupament industrial de noves i més potents substàncies adhesives. No és estrany, doncs, que hi hagi molts estudis al respecte.
Podrà l’ésser humà escalar parets com ho fa l’Spiderman algun dia? Labonte et al. (2016) ens explica per què l’Spiderman com a tal no podria existir. O, almenys, com hauria de ser realment per poder adherir-se a les parets com una aranya.

Sense entrar en les estratègies pròpies de cada organisme (de les quals parlarem després), el principi bàsic pel qual insectes, aranyes o geckos poden caminar sobre superfícies verticals llises o cap per avall és la seva relació superfície/volum: com més petit és l’animal, més gran és la superfície del seu cos respecte al seu volum i menor la quantitat de superfície adhesiva necessària per poder desplaçar-se sense caure a causa del pes. Així doncs, els geckos serien els animals coneguts amb la mida més gran (relació superfície/volum més petita) capaços de caminar sobre superfícies verticals llises o cap per avall sense patir modificacions anatòmiques que farien inviable el seu desenvolupament.
I què vol dir “sense patir modificacions anatòmiques”? Els mateixos autors expliquen que com més gran és l’animal, més gran és la superfície adhesiva necessària per desplaçar-se sense desprendre’s. El creixement de la superfície adhesiva respecte la mida de l’animal segueix un patró d’al·lometria positiva extrema: per un petit increment de la mida de l’animal, es produeix un augment significativament major de la superfície adhesiva. Segons aquest estudi, la superfície adherent respecte a la superfície total pot ser fins a 200 vegades més gran en geckos que en àcars.

No obstant això, la mateixa al·lometria es regeix per una sèrie de constriccions (limitacions) anatòmiques. Així, per tal que existís un animal més gran que un gecko capaç de caminar sobre una superfície vertical llisa o invertida, aquest hauria de desenvolupar, per exemple, unes extremitats enormes amb una superfície adherent igualment gran. Si bé podria tenir sentit des d’un punt de vista físic, les constriccions anatòmiques fan inviable l’existència d’animals amb aquestes característiques.
Ara ja podem respondre la pregunta “Per què l’Spiderman no podria adherir-se a les parets?”. Segons aquest estudi, perquè un ésser humà pogués caminar per les parets com una aranya el seu cos hauria d’estar recobert almenys d’un 40% d’estructures adhesives (un 80% si comptem únicament la seva part frontal); o això, o tenir braços o cames absurdament grans i impossibles des d’un punt de vista anatòmic.
Gran diversitat d’estratègies
L’adhesió dinàmica ha de ser prou forta perquè l’animal no caigui estant quiet, però prou feble per poder desenganxar-sense problemes en fer un pas.
Per aconseguir-ho, hi ha diferents estratègies.

1) Adhesió humida
Hi intervè una substància líquida.
Insectes
Els insectes presenten dos sistemes:
Potes amb coixinets llisos: el trobem, per exemple, en formigues, abelles, paneroles i saltamartins. L’últim segment de les seves potes (pretars), les ungles o les tíbies presenten un o diversos coixinets extremadament tous i deformables (com els arolis al pretars). A petita escala, cap superfície és totalment llisa, de manera que aquests coixinets es deformen fins a ocupar tots els seus espais disponibles.

Potes amb coixinets peluts: el trobem en escarabats i mosques, entre d’altres. Els coixinets d’aquests insectes estan densament coberts de petites estructures similars a pèls, les setes, gràcies a les quals el contacte amb la superfície augmenta.

En ambdós casos, intervé un líquid amb una fase hidrofòbica i una altra hidrofílica. Estudis amb formigues han demostrat que les terminacions de les seves potes secreten una fina capa de líquid que incrementa el contacte entre el pretars i la superfície sobre la que caminen, omplint els buits restants i actuant com un adhesiu sota els principis de capil·laritat (tensió superficial) i viscositat.
Si voleu conèixer més a fons aquest mecanisme, no us perdeu aquest increïble vídeo sobre les formigues!:
Granotes arborícoles
Els coixinets dels dits de les granotes arborícoles estan compostos de cèl·lules epitelials columnars separades entre si. Entre elles, nombroses glàndules hi aboquen una substància mucosa. La separació de les cèl·lules permet, d’una banda, que els coixinets es deformin per adaptar-se al terreny i, per altra, que la mucositat circuli entre elles i asseguri l’adhesió. A més a més, en ambients humits (moltes d’aquestes granotes viuen en selves), aquests espais faciliten l’eliminació de l’excés d’aigua que les faria relliscar.

En el següent vídeo, pots apreciar amb més detall les potes d’una de les granotes arborícoles més conegudes:
Les granotes arborícoles presenten un sistema similar al de coixinets llisos dels insectes. De fet, a molts augments les microestructures adhesives en grills i granotes són pràcticament idèntiques. Això va dur Barnes (2007) a considerar l’adhesió humida com una de les més exitoses.

Pòssums
Els estudis més detallats s’han realitzat sobre el pòssum pigmeu acròbata (Acrobates pygmaeus), un petit marsupial de la mida d’un ratolí capaç d’escalar superfícies de vidre fent servir els grans coixinets dels palmells de les seves potes. Aquests coixinets estan compostos de múltiples capes de cèl·lules epitelials esquamoses separades per solcs que en faciliten la deformació i pels quals hi circula la suor, que és el líquid que fan servir per adherir-se.


2) Adhesió seca
No intervenen líquids.
Aranyes i geckos
Tant les aranyes com els geckos es regeixen pel mateix principi d’adhesió: les forces de Van de Waals. A diferència d’insectes, granotes i pòssums, no segreguen líquids adhesius.
Les forces de Van der Waals resulten de la interacció entre molècules o àtoms sense que hi hagi un enllaç químic entre ells, i la seva energia depèn de la distància. Aquestes interaccions apareixen entre els “pèls” o setes dels palmells de les potes dels geckos (les quals estan solcades per plecs, les lamel·les) i les setes de les potes de les aranyes (que estan cobertes de moltes pilositats formant les escòpules), i la superfície sobre la qual caminen.


Estudis recents, però, suggereixen que les interaccions de Van der Waals no serien les grans determinants de l’adhesió en els geckos, sinó les interaccions electrostàtiques (diferent polaritat entre les setes i la superfície), després de comprovar que la seva capacitat adhesiva minvava sobre materials menys energètics, com el tefló.
Sigui com sigui, l’habilitat dels geckos per enfilar-se és impressionant. Si no, mira aquest vídeo del gran David Attenborough:
3) Succió
Ratpenats
Els ratpenats de ventoses (família Thyropteridae), originaris de l’Amèrica Central i del Sud, presenten unes ventoses en forma de disc als seus polzes i al palmell del segon parell de potes que els permeten desplaçar-se sobre superfícies llises. A l’interior d’aquests discos, la pressió es redueix i el ratpenat queda adherit per succió. De fet, un sol disc pot suportar el pes de tot l’animal.

Després de conèxier totes aquestes estratègies, creus que l’Spiderman n’està a l’alçada?
Imatge de portada d’autor desconegut. Font: link.





















































 El verí de molts animals és útil tant pel desenvolupament d'antídots, com en la recerca de analgèsics i altres medicaments. Foto de l’extracció de verí d’un escurçó (Echis carinatus), de
El verí de molts animals és útil tant pel desenvolupament d'antídots, com en la recerca de analgèsics i altres medicaments. Foto de l’extracció de verí d’un escurçó (Echis carinatus), de  Serp llop de l’Índia (Lycodon aulicus), exemple d’ofidi. Foto de
Serp llop de l’Índia (Lycodon aulicus), exemple d’ofidi. Foto de  Basilisc marró (Basiliscus vittatus), exemple d’iguani. Foto de
Basilisc marró (Basiliscus vittatus), exemple d’iguani. Foto de  Varà sord de Borneo (Lanthanotus borneensis), exemple d’anguimorf. Foto de
Varà sord de Borneo (Lanthanotus borneensis), exemple d’anguimorf. Foto de  Crani d’helodermàtid, en el que observem les esmolades dents amb les que injecten el verí. Imatge de
Crani d’helodermàtid, en el que observem les esmolades dents amb les que injecten el verí. Imatge de  Foto de
Foto de  Foto d’un drac barbut de l’est, en que veiem el interior de la seva boca de color groc. Ens estarà intentant avisar d’algo amb aquesta coloració? Foto de
Foto d’un drac barbut de l’est, en que veiem el interior de la seva boca de color groc. Ens estarà intentant avisar d’algo amb aquesta coloració? Foto de  Secció transversal de la boca d’un drac barbut de l’est, on es veuen les glàndules verinoses incipients tant de la mandíbula superior (mxivg) com de la inferior (mnivg). Imatge extreta de
Secció transversal de la boca d’un drac barbut de l’est, on es veuen les glàndules verinoses incipients tant de la mandíbula superior (mxivg) com de la inferior (mnivg). Imatge extreta de  Varà gegant australià o “perentie” (Varanus giganteus) un varànid típic, amb coll llarg, potes robustes, metabolisme actiu i sentits desenvolupats. Foto de
Varà gegant australià o “perentie” (Varanus giganteus) un varànid típic, amb coll llarg, potes robustes, metabolisme actiu i sentits desenvolupats. Foto de  Foto on veiem la temuda saliva dels varans, concretament d’un varà aquàtic (Varanus salvator). Imatge de
Foto on veiem la temuda saliva dels varans, concretament d’un varà aquàtic (Varanus salvator). Imatge de  Crani de megalània (Varanus priscus) en el que veiem les dents sense solcs. Aquest varà extint de més de 5 metres de llarg, fou l’animal verinós més gran conegut. Foto de
Crani de megalània (Varanus priscus) en el que veiem les dents sense solcs. Aquest varà extint de més de 5 metres de llarg, fou l’animal verinós més gran conegut. Foto de  Grup de dragons de Komodo (Varanus komodoensis) devorant un porc recent caçat. Imatge extreta de
Grup de dragons de Komodo (Varanus komodoensis) devorant un porc recent caçat. Imatge extreta de 

 Imatge de microscopi d’un tall transversal del crani d’una serp, on es veu l’epitel·li olfactiu tant de la cavitat nasal com de l’òrgan vomeronasal. Imatge de Elliott Jacobson.
Imatge de microscopi d’un tall transversal del crani d’una serp, on es veu l’epitel·li olfactiu tant de la cavitat nasal com de l’òrgan vomeronasal. Imatge de Elliott Jacobson. Esquema de la posició de l’òrgan vomeronasal. Aquest es forma durant el desenvolupament embrionari a partir de la cavitat nasal i té una obertura al paladar. Imatge de
Esquema de la posició de l’òrgan vomeronasal. Aquest es forma durant el desenvolupament embrionari a partir de la cavitat nasal i té una obertura al paladar. Imatge de  Els varans (parents de les serps) també presenten una llengua bífida que els permet ensumar l’aire. Foto d’un varà aquàtic (Varanus salvator) a l’Índia, de
Els varans (parents de les serps) també presenten una llengua bífida que els permet ensumar l’aire. Foto d’un varà aquàtic (Varanus salvator) a l’Índia, de  Foto d’una
Foto d’una  Retrat d’una pitó de l’Índia (Python molurus) en el que es veu clarament l’absència d’oïda externa. Foto de
Retrat d’una pitó de l’Índia (Python molurus) en el que es veu clarament l’absència d’oïda externa. Foto de 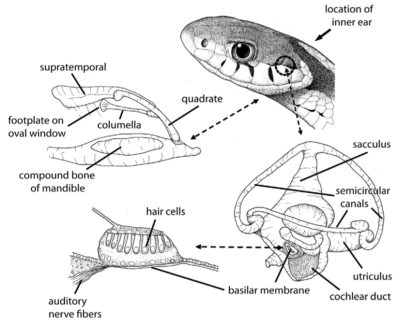 Esquema de l’aparell auditiu d’una serp qualsevol. Imatge de Dan Dourson.
Esquema de l’aparell auditiu d’una serp qualsevol. Imatge de Dan Dourson. Les serps aquàtiques com l’anaconda (Eunectes murinus) poden detectar amb tot el cos els sons d’animals movent-se sota l’aigua. Foto de
Les serps aquàtiques com l’anaconda (Eunectes murinus) poden detectar amb tot el cos els sons d’animals movent-se sota l’aigua. Foto de  L’escurçó banyut aràbic (Cerastes gasperettii) és una serp que
L’escurçó banyut aràbic (Cerastes gasperettii) és una serp que  Els ofidis més primitius, com aquesta serp cega europea (Typhlops vermicularis), tenen ulls petits i poc desenvolupats. Imatge de
Els ofidis més primitius, com aquesta serp cega europea (Typhlops vermicularis), tenen ulls petits i poc desenvolupats. Imatge de  Serp ratera occidental (Pantherophis obsoletus) a punt de mudar de pell, moment en que l’escama ocular es torna opaca. Foto de
Serp ratera occidental (Pantherophis obsoletus) a punt de mudar de pell, moment en que l’escama ocular es torna opaca. Foto de  Moltes serps arborícoles com aquesta serp fuet verda (Ahaetulla nasuta) presenten pupil·les horitzontals que els permeten augmentar el seu camp de visió, fent que puguin calcular millor les distàncies entre branca i branca. Foto de
Moltes serps arborícoles com aquesta serp fuet verda (Ahaetulla nasuta) presenten pupil·les horitzontals que els permeten augmentar el seu camp de visió, fent que puguin calcular millor les distàncies entre branca i branca. Foto de 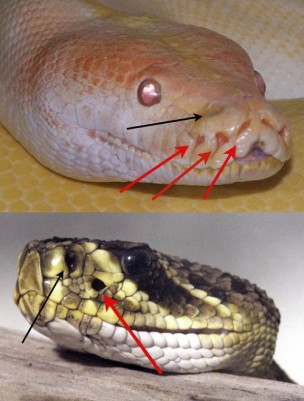 Fotos d’una pitó i un cròtal on es senyalen tant els forats del nas (fletxes negres) com les fossetes termorreceptores (fletxes vermelles). Imatge de
Fotos d’una pitó i un cròtal on es senyalen tant els forats del nas (fletxes negres) com les fossetes termorreceptores (fletxes vermelles). Imatge de  Esquema de l’estructura de la fosseta termorreceptora d’un cròtal. Aquesta presenta una membrana sensible als canvis de temperatura, darrera de la qual hi ha una cambra amb aire i nervis sensibles a l’escalfor. Aquest aire es dilata amb els increments de temperatura i activa el nervi trigèmin. Imatge de
Esquema de l’estructura de la fosseta termorreceptora d’un cròtal. Aquesta presenta una membrana sensible als canvis de temperatura, darrera de la qual hi ha una cambra amb aire i nervis sensibles a l’escalfor. Aquest aire es dilata amb els increments de temperatura i activa el nervi trigèmin. Imatge de 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}