

Desde la prehistoria, donde la medicina tuvo sus comienzos con plantas, minerales y partes de animales; hasta día de hoy, la medicina ha evolucionado a pasos de gigante. Gran parte de la “culpa” de este hecho se la debemos a la genética, que nos permite hablar de medicina personalizada. De este tipo de medicina es de la que trata el siguiente artículo.
LA EVOLUCIÓN DE LAS ENFERMEDADES
Para hablar de medicina debemos conocer primero las enfermedades. Pero no podemos pensar que todas las enfermedades son genéticas, sino que existen enfermedades relacionadas con cambios anatómicos, fruto de nuestra evolución.
El chimpancé es el animal actual más cercano a nosotros, los humanos, con el que compartimos el 99% de nuestro genoma. A pesar de esto, los humanos tenemos características fenotípicas muy particulares como el cerebro más desarrollo, tanto a tamaño como a expansión de la corteza cerebral; piel sudorosa sin pelo, postura bípeda y dependencia prolongada de las crías, que permite la transmisión de conocimientos durante más tiempo; entre otras.
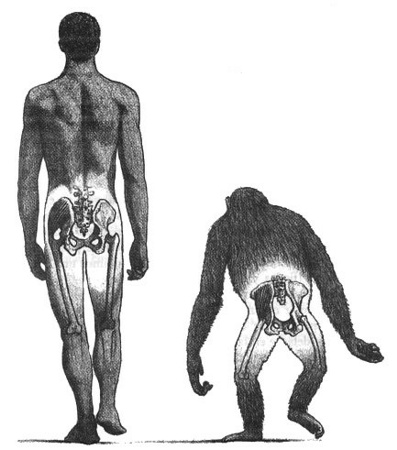
Posiblemente, la postura bípeda fue clave para que se produjera pronto la divergencia entre el linaje de chimpancé y el de humanos; y también es la razón de la aparición de algunas enfermedades relacionadas con factores anatómicos. Entre ellas encontramos hernias, hemorroides, varices, desórdenes de la columna, como hernias de los discos intervertebrales; osteoartritis en la articulación de la rodilla, prolapso uterino y dificultades en el parto.
El hecho de que la pelvis se remodelara (Figura 1) y fuera más estrecha resultó en problemas obstétricos millones de años después, cuando el cerebro se expandió y, por consecuencia, el cráneo también. Las cabezas de los fetos eran más largas y grandes dificultando el parto. Esto explica porque los partos de los humanos son más largos y prolongados en comparación con los de los chimpancés y otros animales.
La evolución hacia la vida moderna nos ha comportado muchos cambios en todos los sentidos. En comparación con nuestros antepasados cazadores y recolectores (Figura 2), nuestra dieta ha cambiado mucho y no tiene nada que ver con lo que comen el resto de primates. Para estos últimos, la fruta representa la mayoría de la ingesta, pero para nosotros lo es la carne roja. Además, somos los únicos animales que seguimos alimentándonos de leche pasado el período de lactancia.

Si al cambio en la dieta le añadimos el sedentarismo y la poca actividad física de los humanos modernos, puede ayudar a explicar la gravedad y frecuencia de algunas enfermedades humanas modernas.
El estilo de vida también puede producirnos afectaciones. Por ejemplo la miopía, que su tasa es mayor en individuos occidentales que leen mucho o hacen actividades de visión cercana, en comparación a individuos de pueblos aborígenes.
Otro ejemplo claro es la alteración en la etapa reproductiva femenina. Actualmente las mujeres tienen hijos cada vez más tarde. Esto también va ligado a una disminución de la duración de la lactancia materna. Estos cambios, que socialmente se pueden considerar positivos, tienen efectos negativos sobre la salud de los órganos reproductivos. Está demostrado que la combinación de menarquia precoz, la lactancia limitada o inexistente y una menopausia más tardía son los principales factores de riesgo para cáncer de mama y ovario.
Los seres humanos cada vez vivimos más años y queremos la mejor calidad de vida. Es fácil que a mayor longevidad aparezcan más enfermedades, por el deterioro del organismo y de sus células.
LA EVOLUCIÓN DE LA MEDICINA
La historia de la medicina es la historia de la lucha de los hombres contra la enfermedad y, desde comienzos de este siglo, también es la historia del esfuerzo humano por mantener la salud.
Los conocimientos científicos de la medicina los hemos adquirido basándonos en la observación y en la experiencia, pero no siempre ha sido así. Nuestros antepasados experimentaron las enfermedades y el temor a la muerte antes de poderse hacer una imagen racional de ellas, y la medicina de entonces se hallaba inmersa en un sistema de creencias, mitos y ritos.
Pero en los últimos años ha nacido la genómica personalizada, que te dice tus factores de riesgo. Esto abre una puerta a la medicina personalizada, que ajusta los tratamientos a los pacientes dependiendo de su genoma (Figura 3). Utiliza la información de los genes y proteínas de una persona para prevenir, diagnosticar y tratar una enfermedad, y todo gracias a la secuenciación del genoma humano.

Los métodos moleculares que hacen posible la medicina de precisión, incluyen pruebas de variación de genes, proteínas y nuevos tratamientos dirigidos a mecanismos moleculares. Con los resultados de estas pruebas y tratamientos se puede determinar el estado de la enfermedad, predecir el estado futuro de la misma, la respuesta al medicamento y el tratamiento o, incluso, el papel de los alimentos que ingerimos en determinados momentos, lo que resulta de gran ayuda a los médicos a individualizar el tratamiento de cada paciente.
Para ello tenemos a nuestro alcance la nutrigenética y la nutrigenómica, que al igual que la farmacogenética y la farmacogenómica, ayudan al avance de una medicina cada vez más dirigida. Por lo tanto, estas disciplinas son hoy en día uno de los pilares de la medicina personalizada, ya que supone tratar cada paciente de forma individualizada y a medida.
La evolución hacia la medicina de precisión es personalizada, preventiva, predictiva y participativa. Cada vez hay más acceso a la información y el paciente es más proactivo, adelantándose a los problemas, previniéndolos o estar preparados para enfrentarlos eficientemente.
REFERENCIAS
-
Varki, A. Nothing in medicine makes sense, except in the light of evolution. J Mol Med (2012) 90:481–494
-
Nesse, R. and Williams, C. Evolution and the origins of disease. Sci Am. (1998) 279(5):86-93
- Mackenbach, J. The origins of human disease: a short story on “where diseases come from”. J Epidemiol Community Health. (2006) 60(1): 81–86
- Foto portada: Todos Somos Uno



















































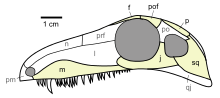 Dibujo de un cráneo de Archaeothyris, en el que podemos ver algunas de las características de los sinápsidos, como la fenestra temporal y dientes caniniformes. Dibujo de
Dibujo de un cráneo de Archaeothyris, en el que podemos ver algunas de las características de los sinápsidos, como la fenestra temporal y dientes caniniformes. Dibujo de 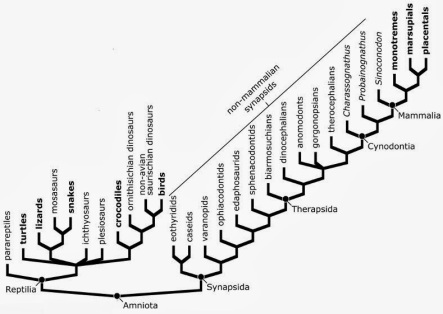 Árbol evolutivo de los amniotas modificado de
Árbol evolutivo de los amniotas modificado de 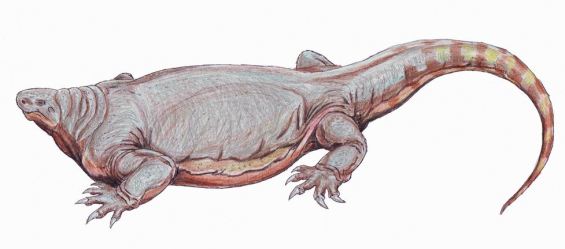 Reconstrucción de Cotylorhynchus, un caseasaurio que crecía hasta los 3 metros de largo. Dibujo de
Reconstrucción de Cotylorhynchus, un caseasaurio que crecía hasta los 3 metros de largo. Dibujo de  Dibujos del varanópido Varanodon (superior) y del ofiacodóntido Ophiacodon (inferior). Dibujos de
Dibujos del varanópido Varanodon (superior) y del ofiacodóntido Ophiacodon (inferior). Dibujos de 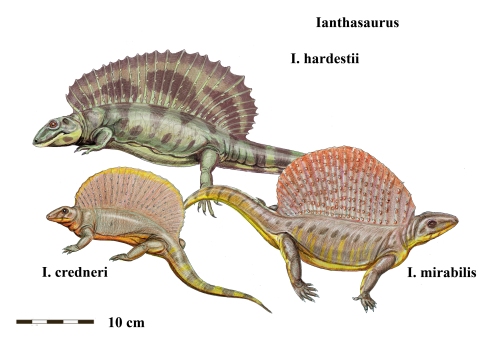 Reconstrucción de diferentes especies de edafosáuridos del género Ianthasaurus, mostrando su vela característica. Dibujos de
Reconstrucción de diferentes especies de edafosáuridos del género Ianthasaurus, mostrando su vela característica. Dibujos de  Esqueleto de Edaphosaurus del Field Museum de Chicago, donde se pueden ver los tubérculos en las espinas neurales. Imagen de
Esqueleto de Edaphosaurus del Field Museum de Chicago, donde se pueden ver los tubérculos en las espinas neurales. Imagen de  Reconstrucción del esfenacodóntido Dimetrodon, de
Reconstrucción del esfenacodóntido Dimetrodon, de  Esqueleto de Biarmosuchus, un terápsido basal en el cual podemos ver su postura más erecta. Imagen de
Esqueleto de Biarmosuchus, un terápsido basal en el cual podemos ver su postura más erecta. Imagen de 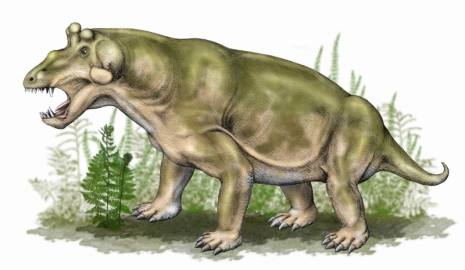 Reconstrucción de Estemmenosuchus, un dinocéfalo del cual se han encontrado impresiones fósiles de la piel, por lo que se sabe que estaba cubierto de piel lisa y glandular sin escamas. Dibujo de
Reconstrucción de Estemmenosuchus, un dinocéfalo del cual se han encontrado impresiones fósiles de la piel, por lo que se sabe que estaba cubierto de piel lisa y glandular sin escamas. Dibujo de 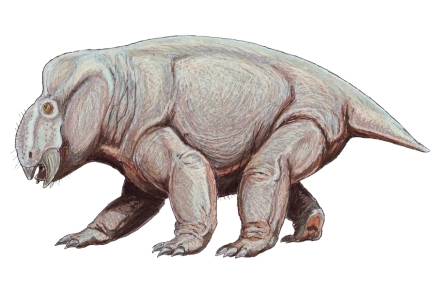 Reconstrucción de Placerias, un anomodonto que podía llegar a pesar hasta una tonelada. Dibujo de
Reconstrucción de Placerias, un anomodonto que podía llegar a pesar hasta una tonelada. Dibujo de 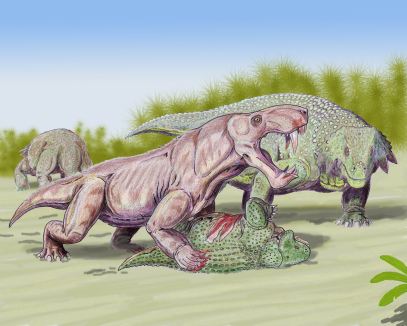 Reconstrucción de Inostrancevia, el género de gorgonópsido más grande, depredando a Scutosaurus, un parareptil. Dibujo de
Reconstrucción de Inostrancevia, el género de gorgonópsido más grande, depredando a Scutosaurus, un parareptil. Dibujo de  Reconstrucción de una pareja de Pristerognathus, un género de terocéfalos en los cuáles podemos ver algunas características de los mamíferos. Dibujo de
Reconstrucción de una pareja de Pristerognathus, un género de terocéfalos en los cuáles podemos ver algunas características de los mamíferos. Dibujo de 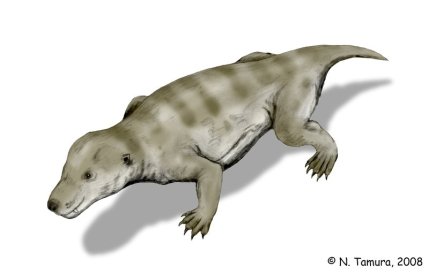

 Reconstrucción de Megazostrodon, un pequeño mamaliaforme que representa bastante bien la transición de cinodontos a mamíferos modernos. Imagen de
Reconstrucción de Megazostrodon, un pequeño mamaliaforme que representa bastante bien la transición de cinodontos a mamíferos modernos. Imagen de 


























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}