
Durant més de 130 anys els dinosaures han estat classificats en dos ordres separats, els saurisquis i els ornitisquis. Però com sempre passa en les ciències biològiques, tota teoria és certa fins que es demostra el contrari. Un nou estudi ha posat en entredit la classificació clàssica dels dinosaures, eliminant i redistribuïnt alguns dels diferents grups de dinosaures. Tot i que aquesta nova hipòtesis no és segura al 100%, en aquesta entrada us explicarem en què consisteix aquesta reordenació dels dinosaures.
CLASSIFICACIÓ TRADICIONAL DELS DINOSAURES
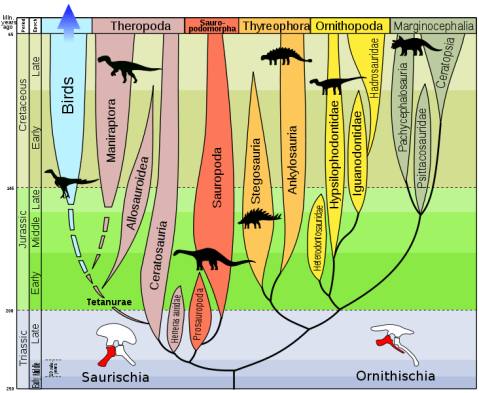
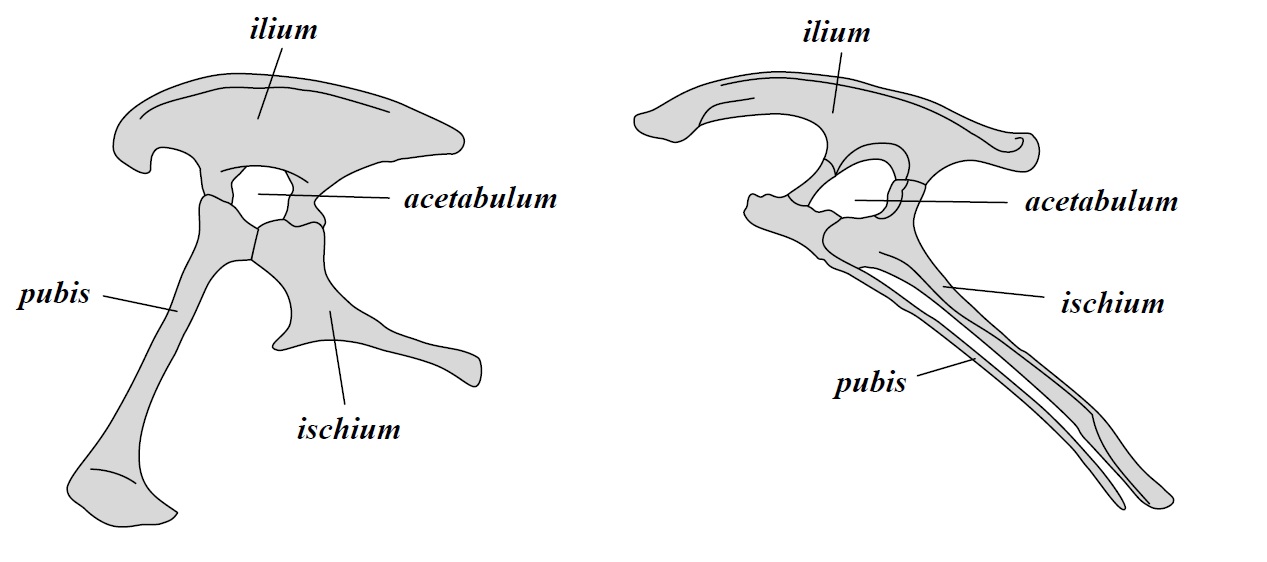
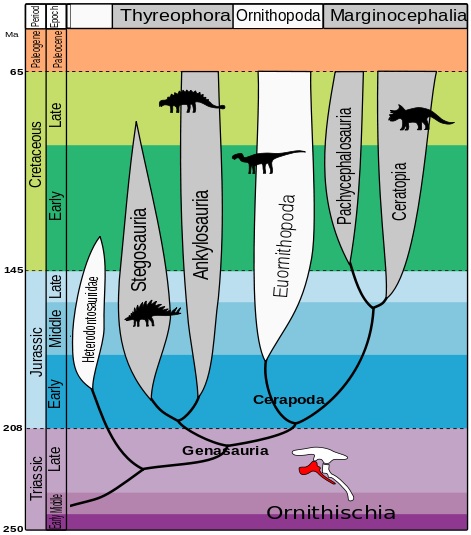
Des del segle XIX, els dinosaures s’han dividit en dos grans ordres basats en la estructura de la seva pelvis. L’ordre Saurischia (pelvis de llangardaix) inclou als teròpodes (dinosaures carnívors i ocells actuals) i als sauropodomorfs (grans herbívors de coll llarg); l’ordre Ornithischia (pelvis d’ocell) inclou als ornitòpodes (dinosaures herbívors i amb bec d’ànec), als marginocèfals (dinosaures amb banyes i estructures cranials endurides) i als tireòfors (dinosaures cuirassats).
Tanmateix, aquesta classificació no té l’última paraula. La paleontologia és una ciència extremadament volàtil, ja que amb cada nou descobriment es pot desmuntar tot el que es sabia fins al moment, encara que es tracti d’una hipòtesi centenària. Això és el que ha passat amb els dinosaures recentment.
L’INICI D’UNA NOVA HIPÒTESI
Un nou estudi publicat el març del 2017, ha fet replantejar la classificació tradicional del dinosaures. Molts estudis anteriors assumien com a certa la classificació en Saurischia/Ornithischia tradicional i per tant, els caràcters i els taxons utilitzats ja anaven enfocats en aquesta classificació. En canvi, aquest nou estudi ha estat pioner en varis aspectes:
- Inclou un gran nombre d’espècies i taxons (molts més que en investigacions anteriors).
- Estudis anteriors donaven molt més ènfasi en dinosaures teròpodes i sauropodomorfs (saurisquis tradicionals) basals, ja que van ser dels primers grups en diversificar-se, incloent pocs ornitisquis basals.
- També s’hi han inclòs molts arcosaures dinosauromorfs (taxons no dinosaures).
- Estudis anteriors havien assumit que molts caràcters dels ornitisquis eren simplesiomòrfics (caràcters ancestrals de tots els dinosaures) i només es centraven en poques sinapomorfies (caràcters compartits per un grup monofilètic).
Aquest estudi s’ha desprès de moltes de les presuposicions anteriors sobre filogènia dinosauria i ha analitzat un gran nombre de espècies i multitud de caràcters no inclosos en investigacions anteriors. Això ha fet que l’arbre evolutiu resultant fos molt diferent dels que s’havien obtingut fins llavors.
RECONSTRUÏNT L’ARBRE
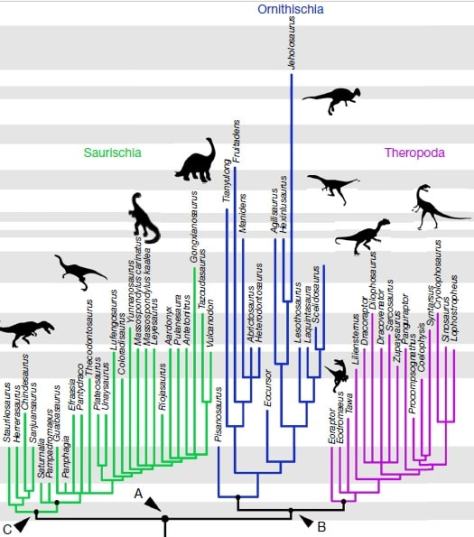
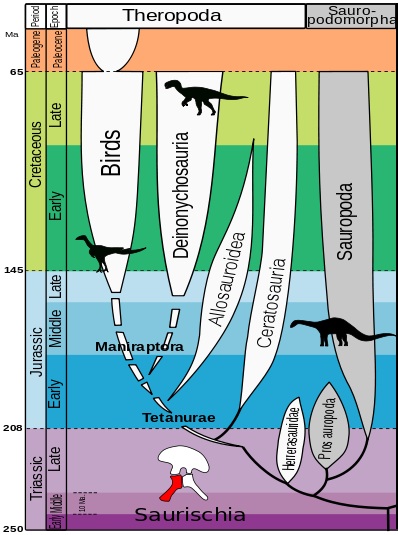
Llavors, com queda l’arbre evolutiu dels dinosaures segons aquesta hipòtesi? Doncs el tema és una mica complicat, tot i que els diferents taxons segueixen quedant dividits en dos ordres:
- L’ordre Saurischia que, segons l’estudi, només inclou als sauropodomorfs i als herrerasàurids (grup de saurisquis carnívors no teròpodes).
- El nou ordre Ornithoscelida (potes d’ocell) que inclou als ornitisquis tradicionals i als teròpodes, que deixen de ser saurisquis.
Tenint això en ment, vegem doncs les característiques que defineixen aquests dos grups.
Saurisquis
L’ordre Saurischia es manté pràcticament igual, excepte que els teròpodes es queden fora del grup. Aquest ordre presenta la pelvis d’estructura saurisquia original, com també la presentaven els avantpassats dels dinosaures. Segons la nova hipòtesi, dins dels saurisquis s’hi inclouen els herrerasàurids i els sauropodomorfs.
Els herrerasàurids (família Herrerasauridae) van ser un petit grup de saurisquis basals que van evolucionar cap a una dieta carnívora. Per això durant un temps es pensava que eren el grup germà dels teròpodes, però després es va veure que el seu lloc es trobava entre els primers saurisquis. Tot i estar força especialitzats, probablement la competència amb altres depredadors va fer que no duressin molt de temps, apareixent a mitjans del Triàssic i extingint-se al finals d’aquest període.

Els herrerasàurids ocupaven un nínxol ecològic similar al dels teròpodes. La nova hipòtesi implica doncs que la hipercanivoria (alimentació exclusivament de carn) va evolucionar independentment dos cops en els dinosaures, cosa que fa que alguns paleontòlegs la posin en dubte. Tanmateix, l’anatomia dels herrerasàurids i dels teròpodes diferia en certs aspectes, com la anatomia de les mans (més generalista en els herrerasàurids) i la estructura de la mandíbula.
Els primers sauropodomorfs eren animals bípedes igual que els herrerasàurids, tot i que aquests eren animals omnívors. Tanmateix, els sauropodomorfs acabarien convertint-se en enormes quadrúpedes herbívors amb colls llarguíssims característics.

Ornitoscèlids
El nou ordre de dinosaures és Ornithoscelida, que agrupa els teròpodes amb els ornitisquis. Aquest taxó està recolzada per més de vint sinapomorfies (caràcters derivats compartits per un clade) esquelètiques, presents tant en teròpodes com en ornitisquis basals. Algunes d’aquestes característiques inclouen la presència d’una separació entre les dents premaxil·lars i maxil·lars (diastema) i la fusió dels extrems de la tíbia i la fíbula en un tibiotars (tot i que aquests caràcters només es troben en les espècies més basals).
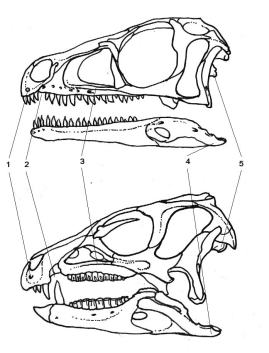
Tant els teròpodes com els primers ornitisquis eren animals bípedes. A més, la presència de dents heterodontes en els membres ancestrals d’ambdós grups fa pensar que els primers ornitoscèlids eren omnívors, els quals després s’especialitzarien en alimentar-se de carn i de vegetació (teròpodes i ornitisquis respectivament).

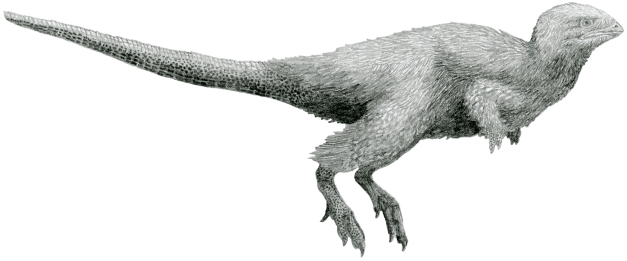
Una curiositat de la nova classificació és que acceptant a Ornithoscelida com un taxó vàlid, s’agrupen a tots els dinosaures amb plomes en un sol grup. Per tots és sabut que molts teròpodes presentaven plomes (ja que eren els avantpassats dels ocells) però, el que molta gent desconeix és que la presència de plomes també s’ha demostrat en alguns ornitisquis basals i en altres més avançats.
SEGUIR INVESTIGANT
I llavors, és definitiva aquesta hipòtesi? Doncs no, per descomptat. Tot i que resulta força atractiu dir que s’ha canviat la història natural dels dinosaures per sempre, no podem dir que a partir d’ara els dinosaures es classifiquin d’aquesta manera.
Encara que aquest estudi mostri resultats interessants sobre l’origen dels dinosaures, no es poden ignorar els centenars d’estudis anteriors que s’han fet sobre aquest grup d’animals. Haurem d’estar atents a nous articles que poc a poc vagin desvelant més informació sobre les relacions entre aquests rèptils mesozoics. I és que això és el més estimulant de la biologia, que no hi ha res segur! I que amb noves tècniques d’estudi i nous descobriments, poc a poc aprenem més sobre el món que ens envolta.
Mantingueu la ment oberta i seguiu investigant!
REFERÈNCIES
S’han consultat les següents fonts durant l’elaboració d’aquesta entrada:
- G. Seeley (1887). On the Classification of the Fossil Animals Commonly Named Dinosauria. Proceedings of the Royal Society of London Vol. 43. Pp 165-171.
- Baron, Norman & Barret (2017). A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature Vol. 543. Pp 501-506.
- Senter & Robins (2015). Resting Orientations of Dinosaur Scapulae and Forelimbs: A Numerical Analysis, with Implications for Reconstructions and Museum Mounts. Plos One.
- Scientific American. Ornithoscelida Rises: A New Family Tree for Dinosaurs.
- Imatge de portada de Charles Knight.













































{kind=link}
{kind=link}