

Turtles are charming animals yet, while they look cute to most people, they’ve been racking the brains of palaeontologists for decades. The combination of apparently primitive features and an extremely specialized anatomy, has made the reconstruction of the origin and evolution of these reptiles a nearly impossible task. In this entry we’ll try to get a general idea about the evolution of one of the most striking characteristics of turtles (the shell) with only four examples of primitive “turtles”.
CURRENT AND EXTINCT RELATIVES
As we explained in an earlier entry, the origin of turtles is still debated among the scientific community. Turtles show some anatomic characteristics not found among any current vertebrate, which makes their phylogenetic origin confusing. One of the characteristics that has puzzled palaeontologist more is their skull.
While the rest of reptiles are diapsid (they present a pair of temporal openings at each side of the skull), turtles present a typically anapsid cranium (without any temporal openings). Yet, recent genomic studies have proved that it’s more likely that testudines (order Testudines, current turtles) descend from a diapsid ancestor and that through their evolution they reverted back to the primitive anapsid form. What is not so clear is if turtles are more closely related to lepidosaurs (lizards, snakes and tuataras) or to archosaurs (crocodiles and birds). The most accepted hypothesis is the second one.
Even if the origins of the testudines are still somewhat mysterious, most palaeontologists coincide in that they belong to the clade Pantestudines, which groups all those species more closely related to turtles than to any other animal. A group of reptiles that are also found inside the pantestudines are the sauropterygians like plesiosaurs and placodonts.

EVOLUTION OF TESTUDINES
The rest of pantestudines help us to form an image of how turtles acquired such a specialized anatomy. But first, take a look at some of the characteristics of turtles:
- A shell made up of two parts: the upper shell (carapace) which comes from the fusion of the vertebrae and the dorsal ribs and the lower shell (plastron) that originates from ventral ribs called “gastralia” (still present in some current reptiles).
- While the rest of vertebrates present the scapula over their ribs, the turtle’s ribs (their carapace) cover the scapula.
- The ability to hide their heads and limbs in their shells.
- The absence of teeth; having instead horny ridges in their jaws.
As we’ll see, these characteristics were acquired very gradually.

Even if their relationship with turtles isn’t still very clear, Eunotosaurus africanus is the most ancient candidate to being a turtle’s relative. Eunotosaurus was a fossorial animal that lived 260 million years ago in South Africa. This animal had very wide dorsal ribs which contacted each other, which is thought to have served as an anchoring point for powerful leg muscles, used while digging. Also, similarly to current turtles, Eunotosaurus had lost the intercostal muscles and presented a reorganization of the respiratory musculature.

The oldest indisputable relative of turtles is Pappochelys rosiane from Germany (240 million years ago). The name “Pappochelys” literally means “grandfather turtle” as, before the discovery of Eunotosaurus it was the oldest turtle relative. Just like Eunotosaurus, it presented wide dorsal ribs in contact with each other. Also, its ventral ribs were already wider and thicker and its scapular girdle was placed below the dorsal ribs.
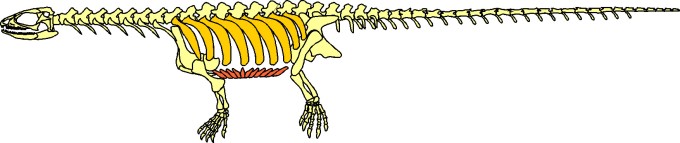
The next step in the evolution of turtles is found 220 million years ago, during the late Triassic in China. Its name is Odontochely semitestacea, which means “toothed turtle with half a shell”. This name is due to the fact that, unlike true turtles, Odontochelys still had a mouth full of teeth and it only presented the lower half of the shell, the plastron. Even if it also had thick dorsal ribs, only paleontological proofs of the plastron have been found. Odontochelys was discovered in freshwater deposits, leads us to believe that at first it only developed the plastron to protect itself from predators attacking from below.

The first testudine known to possess a complete shell is Proganochelys quenstedti from the Triassic period, 210 million years ago. It already presented many characteristics found in current turtles: the shell was completely formed, with carapace and plastron, its skull was anapsid looking and it had no teeth. However, Proganochelys wasn’t able to retract its head and legs inside its shell (even if this may be because of the horns it had). It also presented two extra shell pieces at both sides, which probably served to protect its legs.

PRESENT DAY TURTLES
The order Testudines as we know it, appeared around 190 million years ago, during the Jurassic period. These current turtles are classified into two different suborders, which both separated quickly at the beginning of the evolution of testudines:
Suborder Pleurodira: This suborder is the smallest one as it only contains three current families, all native from the southern hemisphere. The main characteristic is the form in which they retract their neck laterally inside their shell, which leaves the neck exposed and makes the cervical vertebrae present a characteristic shape (Pleurodira roughly means “side neck”). Also, pleurodirans present 13 scutes in their plastrons.

Suborder Cryptodira: Cryptodirans comprise most turtles. While pleurodirans only include freshwater species (as the testudines common ancestor is thought to be), criptodirans include freshwater terrapins, terrestrial tortoises and sea turtles. Apart from only presenting between 11 and 12 scutes in their plastrons, their principal characteristic is the ability to retract their neck and to hide their heads completely in their shell (Cryptodira roughly means “hidden neck”). Cryptodirans are found in practically all the continents and oceans (except in the coldest habitats).

Even if there still are some questions to be answered about the evolution of turtles, we hope that with this little introduction to some of the most characteristic fossil “turtles”, you have had an overall view about how turtles got their shells. Whatever their origins are, we hope that the apparition of men isn’t what puts an end to the history of this group of slow but steady creatures.
REFERENCES
The following sources have been consulted during the elaboration of this entry:
- Bever, Lyson, Field & Bhullar (2015). Evolutionary origin of the turtle skull. Nature. Vol. 525. Pp: 239-242.
- Phenomena, National Geographic. How the turtle got its shell through skeletal shifts and muscular origami.
- Lyson, Bever, Scheyer, Hsiang & Gauthier (2013). Evolutionary Origin of the Turtle Shell. Current Biology. Vol. 23. Pp: 1113-1119.
- Lyson, Rubidge, Scheyer, Queiroz, Schchner, Smith, Botha-Brink & Bever (2016). Fossorial Origin of the Turtle Shell. Current Biology. Vol. 26. Pp: 1887-1894.
- The Atlantic. Why Turtles Evolved Shells: It Wasn’t for Protection.
- Schoch & Sues (2015). A Middle Triassic stem-turtle and the evolution of the turtle body plan. Nature. Vol. 523. Pp: 584-587.
- Reptile Evolution. Odontochelys.
- Cover photo by Daderot.









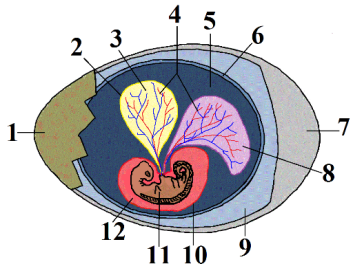 Esquema de l’ou d’un cocodril: 1. closca de l’ou 2. sac vitel·lí 3. vitel (nutrients) 4. vasos sanguinis 5. amni 6. cori 7. aire 8. alantoide 9. albúmina (clara de l’ou) 10. sac amniòtic 11. embrió 12. líquid amniòtic. Imatge de
Esquema de l’ou d’un cocodril: 1. closca de l’ou 2. sac vitel·lí 3. vitel (nutrients) 4. vasos sanguinis 5. amni 6. cori 7. aire 8. alantoide 9. albúmina (clara de l’ou) 10. sac amniòtic 11. embrió 12. líquid amniòtic. Imatge de 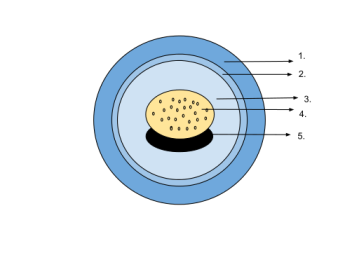 Esquema de l’ou d’un amfibi: 1. càpsula gelatinosa 2. membrana vitel·lina 3. fluid perivitel·lí 4. vitel 5. embrió. Imatge de
Esquema de l’ou d’un amfibi: 1. càpsula gelatinosa 2. membrana vitel·lina 3. fluid perivitel·lí 4. vitel 5. embrió. Imatge de  Reconstrucció de Solenodonsaurus janenschi, un dels candidats a ser el primer amniota, que visqué fa 320-305 milions d’anys a l’actual República Txeca. Recontrucció de
Reconstrucció de Solenodonsaurus janenschi, un dels candidats a ser el primer amniota, que visqué fa 320-305 milions d’anys a l’actual República Txeca. Recontrucció de 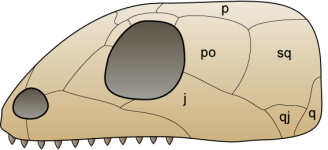 Esquema d’un crani anàpsid, de
Esquema d’un crani anàpsid, de 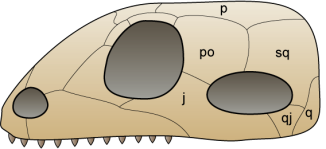 Esquema d’un crani sinàpsid, de
Esquema d’un crani sinàpsid, de 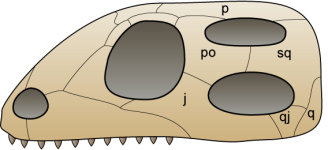 Esquema d’un crani diàpsid, de
Esquema d’un crani diàpsid, de 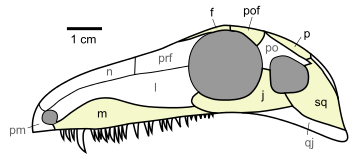 Dibuix del crani de Archaeothyris, el que es creu que va ser un dels primers sinàpsids que visqué fa uns 306 milions d’anys a Nova Escòcia. Dibuix de
Dibuix del crani de Archaeothyris, el que es creu que va ser un dels primers sinàpsids que visqué fa uns 306 milions d’anys a Nova Escòcia. Dibuix de  Reconstrucció de Dimetrodon grandis, un dels sinàpsids més coneguts, de fa uns 280 milions d’anys. Reconstrucció de
Reconstrucció de Dimetrodon grandis, un dels sinàpsids més coneguts, de fa uns 280 milions d’anys. Reconstrucció de 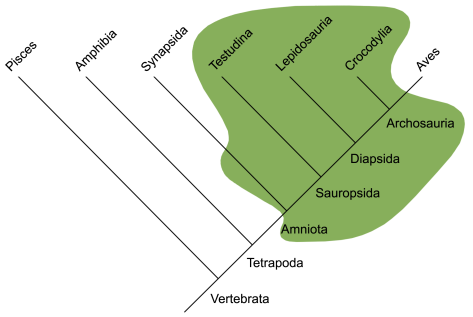 Arbre evolutiu dels vertebrats actuals, on es marca de color verd als grups antigament considerats rèptils. Com es veu, la concepció tradicional de “rèptil” inclou als avantpassats dels mamífers i exclou a les aus. Imatge de
Arbre evolutiu dels vertebrats actuals, on es marca de color verd als grups antigament considerats rèptils. Com es veu, la concepció tradicional de “rèptil” inclou als avantpassats dels mamífers i exclou a les aus. Imatge de  Muda sencera de la pell d’una serp rata. Foto de
Muda sencera de la pell d’una serp rata. Foto de  Foto d’una tuatara (Sphenodon punctatus), de
Foto d’una tuatara (Sphenodon punctatus), de  Fotos d’alguns escamosos d’esquerra a dreta i de dalt a baix: Iguana verda (Iguana iguana, de
Fotos d’alguns escamosos d’esquerra a dreta i de dalt a baix: Iguana verda (Iguana iguana, de 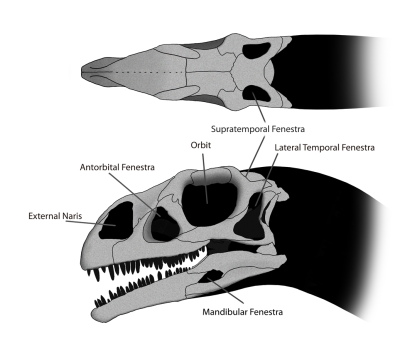 Dibuix del crani del dinosaure Massospondylus en el que es veuen les diferents obertures que caracteritzen als arcosaures diàpsids. Imatge de
Dibuix del crani del dinosaure Massospondylus en el que es veuen les diferents obertures que caracteritzen als arcosaures diàpsids. Imatge de  Foto de dues espècies d’arcosaures actuals; un cocodril del Nil (Crocodylus niloticus) i un tàntal africà (Mycteria ibis). Foto de
Foto de dues espècies d’arcosaures actuals; un cocodril del Nil (Crocodylus niloticus) i un tàntal africà (Mycteria ibis). Foto de  Esquelet de la tortuga extingida Meiolania platyceps que visqué a Nova Caledònia fins fa 3000 anys. En aquesta foto s’aprecia el crani compacte i sense obertures temporals. Foto de
Esquelet de la tortuga extingida Meiolania platyceps que visqué a Nova Caledònia fins fa 3000 anys. En aquesta foto s’aprecia el crani compacte i sense obertures temporals. Foto de  Exemplar de tortuga lleopard (Stigmochelys pardalis) de Tanzània. Foto de
Exemplar de tortuga lleopard (Stigmochelys pardalis) de Tanzània. Foto de 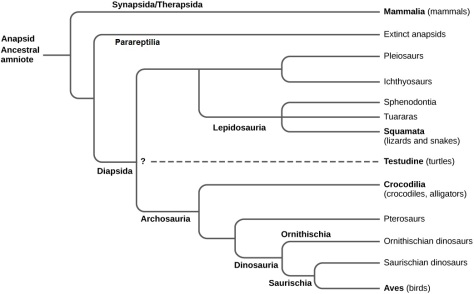 Esquema
Esquema 



{kind=link}
{kind=link}
{kind=link}
{kind=link}