

Els deserts són uns dels hàbitats més extrems del planeta. El del Sàhara, el del Gobi i el de Sonora són exemples d’alguns dels deserts càlids on les altes temperatures i la falta d’aigua suposen un gran repte pels animals que hi viuen. Els rèptils són un dels grups d’animals que presenten les adaptacions més increïbles per a la vida al desert. En aquesta entrada us explicarem els problemes als que s’enfronten els rèptils que hi viuen, i us presentarem diferents espècies de serps i llangardaixos que han trobat en el desert la seva llar.
ELS RÈPTILS AL DESERT
La característica que uneix a tots els deserts és l’escassa precipitació ja que, contràriament al que molta gent pensa, no tots els deserts presenten temperatures altes (existeixen també els deserts freds, com el desert Àrtic i l’Antàrtic, ambdós en perill pel canvi climàtic). Els rèptils són més abundants en els deserts càlids que en els freds, ja que les baixes temperatures els impedirien dur a terme la seva activitat vital.

Els deserts càlids no sempre tenen temperatures extremadament altes. Mentre que durant el dia les temperatures poden arribar a sobrepassar els 45°C, quan es pon el sol les temperatures poden descendir fins a sota del punt de congelació, creant oscil·lacions diàries de fins a 22°C. Els diferents rèptils del desert, al ser poiquiloterms i ectoterms, utilitzen diferents estratègies comportamentals per tal d’evitar el sobreescalfament durant el dia i conservar la temperatura durant la nit (per exemple, enfilant-se a zones elevades o dormint en caus).
El camaleó de Namaqua (Chamaleo namaquensis) regula la seva temperatura corporal canviant de color. A les primeres hores de sol és de color negre per absorbir el màxim de radiació i activar el seu metabolisme. Quan les temperatures augmenten massa, es torna de color blanc per a reflectir la radiació solar. Vídeo de la BBC.
Com ja hem dit, la principal característica de qualsevol desert és la manca d’aigua. En general, en un desert cauen menys de 250 mm d’aigua a l’any. La pell escamosa i impermeable dels rèptils evita la pèrdua d’aigua i els seus excrements contenen àcid úric que, comparat amb la urea, és molt menys soluble en l’aigua, fent que retinguin més líquids. La majoria de rèptils dels deserts extrauen l’aigua que necessiten de l’aliment i alguns beuen l’aigua de la rosada.
Tant les temperatures extremes com les poques precipitacions fan que en els deserts generalment hi hagi poca vida. La vegetació és escassa i els animals solen ser petits i discrets. Aquesta manca de recursos fa que els rèptils del desert siguin més aviat petits comparats amb els seus parents d’ambient més benèvols. A més aquests sauris solen ser animals que aprofiten qualsevol aliment disponible, tot i que s’ho pensen molt bé a l’hora de gastar la seva valuosa energia per aconseguir el seu següent àpat.
SERPS DE LA SORRA
En molts deserts sorrencs hi trobem vàries espècies de serps (i de llangardaixos àpodes) que s’han adaptat a la vida entre les dunes. Molts d’aquests ofidis comparteixen un mètode de desplaçament anomenat “a cops laterals” (en anglès “sidewinding”), en el qual aixequen el cap i coll de terra i els mouen lateralment, mentre que la resta del cos es queda a terra. Quan tornen a posar el cap a terra, el cos s’aixeca fent que les serps es desplacin lateralment en un angle de 45°. Aquest mètode de locomoció fa que les serps dels deserts es desplacin de forma molt eficaç en un terreny inestable. A més, també minimitza el contacte amb un substrat extremament calent, ja que el cos d’aquests ofidis només toca el terra en dos punts en tot moment.
Com veiem en aquest vídeo de RoyalPanthera, el “sidewinding” permet a les serps del desert desplaçar-se minimitzant el contacte amb el terra calent.
Molts ofidis del desert s’enterren a la sorra tant per a evitar la insolació com per a camuflar-se i sorpendre a les seves preses. Això ha fet que moltes serps desertícoles siguin sensibles a les vibracions generades per les seves preses al moure’s per la sorra. A més algunes espècies presenten l’escama rostral (l’escama de la punta del musell) més engruixida i desenvolupada per a ajudar-les a excavar en terrenys sorrencs.

Els escurçons banyuts del gènere Cerastes també presenten vàries característiques que els faciliten la vida als deserts. Aquests escurçons eviten les altes temperatures sent actius durant la nit i passen el dia enterrats a la sorra. El seu mètode de caça consisteix en enterrar-se esperant a que passi una presa, estalviant així el màxim d’energia. Les seves escames supraoculars en forma de banya es creu que els serveixen per a evitar que la sorra cobreixi els seus ulls quan estàn enterrats.

CRIATURES ESPINOSES
En diferents deserts del món hi trobem rèptils que tenen el cos recobert d’espines. Això, no només els proporciona certa protección contra els depredadors, sinó que a més els camufla en un ambient on abunden les plantes espinoses. Dos d’aquests animals són membres del subordre Iguania: el diable espinós i els llangardaixos cornuts.

El diable espinós (Moloch horridus) és un agàmid que viu en deserts sorrencs d’Austràlia. Aquest llangardaix presenta espines per tot el cos que el fan difícil d’empassar per als seus depredadors. També presenta una protuberància darrera del cap que actua com a magatzem de greix. Quan es sent amenaçat, amaga el seu cap autèntic entre les potes i mostra la protuberància del coll com un cap fals. Probablement l’adaptació més interessant d’aquest animal és el sistema de petits canals que presenta entre les escames, els quals recullen tota aigua que entra en contacte amb la pell i la condueixen directament a la boca.
Els llangardaixos cornuts (gènere Phrynosoma, coneguts també com a “gripaus banyuts”) són iguànids que es troben en diferents hàbitats àrids d’Amèrica del Nord. De forma similar al diable espinós, els seus cossos recoberts d’espines els fan difícils d’empassar pels depredadors. A més, al ser atrapats inflen el seu cos per dificultar’ls-hi encara més la tasca. Finalment, algunes espècies com el llangardaix cornut de Texas (Phrynosoma cornutum) són coneguts per la seva capacitat d’autohemorragia: quan es veuen acorralats poden ejectar un raig de sang pudent de l’ull que espanta a la majoria de depredadors.

Com podeu veure, en els deserts hi podem trobar rèptils amb algunes de les adaptacions més enginyoses (i fastigoses) del món. Aquests només són uns pocs exemples de la increïble diversitat de sauris que trobem pels deserts del món, els quals només procuren sobreviure a les dures condicions d’aquests ambients tant extrems. A vegades però, només cal evitar cremar-se els peus amb la sorra.
Vídeo de BBCWorldwide d’un llangardaix musell de pala (Zeros anchietae) fent la “dansa termal” per disminuir el contacte amb la sorra calenta.
REFERÈNCIES
S’han consultat les següents fonts durant l’elaboració d’aquesta entrada:
- Halliday & Adler (2007). La gran enciclopedia de los Anfibios y Reptiles. Editorial Libsa.
- Digital-Desert. Desert Reptiles.
- Arizona-Sonora Desert Museum. Adaptations of Desert Amphibians & Reptiles.
- In the desert. A comprehensive list of the venomous snakes found in the desert.
- H.E.R.P. Herpetological Education & Research Project.
- Christopher J. Bell, Jim I. Mead & Sandra L. Swift (2009). Cranial osteology of Moloch horridus (Reptilia: Squamata: Agamidae). Records of the Western Australian Museum. Vol 25. Pp: 201-237.
- Horned Lizard Conservation Society.
- Imatge de portada de Yathin S. Krishnappa.








 El veneno de muchos animales es útil tanto para el desarrollo de antídotos, como para la investigación de analgésicos y otros medicamentos. Foto de la extracción de veneno de una víbora gariba (Echis carinatus), de
El veneno de muchos animales es útil tanto para el desarrollo de antídotos, como para la investigación de analgésicos y otros medicamentos. Foto de la extracción de veneno de una víbora gariba (Echis carinatus), de  Serpiente lobo de la India (Lycodon aulicus), ejemplo de ofidio. Foto de
Serpiente lobo de la India (Lycodon aulicus), ejemplo de ofidio. Foto de  Basilisco marrón (Basiliscus vittatus), ejemplo de iguanio. Foto de
Basilisco marrón (Basiliscus vittatus), ejemplo de iguanio. Foto de  Varano sordo de Borneo (Lanthanotus borneensis), ejemplo de anguimorfo. Foto de
Varano sordo de Borneo (Lanthanotus borneensis), ejemplo de anguimorfo. Foto de 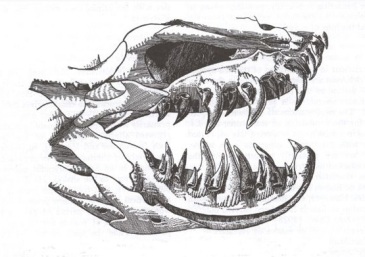 Cráneo de helodermátido, en el que observamos los afilados dientes con los que inyectan el veneno. Imagen de
Cráneo de helodermátido, en el que observamos los afilados dientes con los que inyectan el veneno. Imagen de  Foto de
Foto de  Foto de un dragón barbudo del este, en la que vemos el interior de su boca de color amarillo. ¿Nos estará intentando avisar de algo con esta coloración? Foto de
Foto de un dragón barbudo del este, en la que vemos el interior de su boca de color amarillo. ¿Nos estará intentando avisar de algo con esta coloración? Foto de 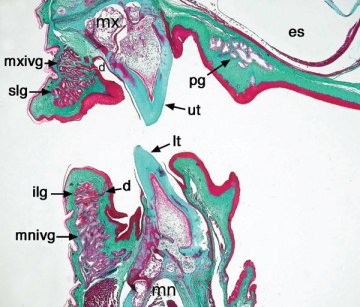 Sección transversal de la boca de un dragón barbudo del este, donde se ven las glándulas venenosas incipientes tanto de la mandíbula superior (mxivg) como de la inferior (mnivg). Imagen extraída de
Sección transversal de la boca de un dragón barbudo del este, donde se ven las glándulas venenosas incipientes tanto de la mandíbula superior (mxivg) como de la inferior (mnivg). Imagen extraída de  Varano gigante australiano o “perentie” (Varanus giganteus) un varánido típico, con cuello largo, patas robusta, metabolismo activo y sentidos desarrollados. Foto de
Varano gigante australiano o “perentie” (Varanus giganteus) un varánido típico, con cuello largo, patas robusta, metabolismo activo y sentidos desarrollados. Foto de  Foto donde vemos la temida saliva de los varanos, concretamente de un varano acuático (Varanus salvator). Imagen de
Foto donde vemos la temida saliva de los varanos, concretamente de un varano acuático (Varanus salvator). Imagen de  Cráneo de megalania (Varanus priscus) en el que vemos los dientes sin surcos. Este varano extinto de más de 5 metros de largo, fue el animal venenoso más grande conocido.
Cráneo de megalania (Varanus priscus) en el que vemos los dientes sin surcos. Este varano extinto de más de 5 metros de largo, fue el animal venenoso más grande conocido.  Grupo de varanos de Komodo (Varanus komodoensis) devorando un cerdo recién cazado. Imagen extraída de
Grupo de varanos de Komodo (Varanus komodoensis) devorando un cerdo recién cazado. Imagen extraída de 



 Esquema de l’ou d’un cocodril: 1. closca de l’ou 2. sac vitel·lí 3. vitel (nutrients) 4. vasos sanguinis 5. amni 6. cori 7. aire 8. alantoide 9. albúmina (clara de l’ou) 10. sac amniòtic 11. embrió 12. líquid amniòtic. Imatge de
Esquema de l’ou d’un cocodril: 1. closca de l’ou 2. sac vitel·lí 3. vitel (nutrients) 4. vasos sanguinis 5. amni 6. cori 7. aire 8. alantoide 9. albúmina (clara de l’ou) 10. sac amniòtic 11. embrió 12. líquid amniòtic. Imatge de  Esquema de l’ou d’un amfibi: 1. càpsula gelatinosa 2. membrana vitel·lina 3. fluid perivitel·lí 4. vitel 5. embrió. Imatge de
Esquema de l’ou d’un amfibi: 1. càpsula gelatinosa 2. membrana vitel·lina 3. fluid perivitel·lí 4. vitel 5. embrió. Imatge de  Reconstrucció de Solenodonsaurus janenschi, un dels candidats a ser el primer amniota, que visqué fa 320-305 milions d’anys a l’actual República Txeca. Recontrucció de
Reconstrucció de Solenodonsaurus janenschi, un dels candidats a ser el primer amniota, que visqué fa 320-305 milions d’anys a l’actual República Txeca. Recontrucció de  Esquema d’un crani anàpsid, de
Esquema d’un crani anàpsid, de  Esquema d’un crani sinàpsid, de
Esquema d’un crani sinàpsid, de  Esquema d’un crani diàpsid, de
Esquema d’un crani diàpsid, de  Dibuix del crani de Archaeothyris, el que es creu que va ser un dels primers sinàpsids que visqué fa uns 306 milions d’anys a Nova Escòcia. Dibuix de
Dibuix del crani de Archaeothyris, el que es creu que va ser un dels primers sinàpsids que visqué fa uns 306 milions d’anys a Nova Escòcia. Dibuix de  Reconstrucció de Dimetrodon grandis, un dels sinàpsids més coneguts, de fa uns 280 milions d’anys. Reconstrucció de
Reconstrucció de Dimetrodon grandis, un dels sinàpsids més coneguts, de fa uns 280 milions d’anys. Reconstrucció de  Arbre evolutiu dels vertebrats actuals, on es marca de color verd als grups antigament considerats rèptils. Com es veu, la concepció tradicional de “rèptil” inclou als avantpassats dels mamífers i exclou a les aus. Imatge de
Arbre evolutiu dels vertebrats actuals, on es marca de color verd als grups antigament considerats rèptils. Com es veu, la concepció tradicional de “rèptil” inclou als avantpassats dels mamífers i exclou a les aus. Imatge de  Muda sencera de la pell d’una serp rata. Foto de
Muda sencera de la pell d’una serp rata. Foto de  Foto d’una tuatara (Sphenodon punctatus), de
Foto d’una tuatara (Sphenodon punctatus), de  Fotos d’alguns escamosos d’esquerra a dreta i de dalt a baix: Iguana verda (Iguana iguana, de
Fotos d’alguns escamosos d’esquerra a dreta i de dalt a baix: Iguana verda (Iguana iguana, de  Dibuix del crani del dinosaure Massospondylus en el que es veuen les diferents obertures que caracteritzen als arcosaures diàpsids. Imatge de
Dibuix del crani del dinosaure Massospondylus en el que es veuen les diferents obertures que caracteritzen als arcosaures diàpsids. Imatge de  Foto de dues espècies d’arcosaures actuals; un cocodril del Nil (Crocodylus niloticus) i un tàntal africà (Mycteria ibis). Foto de
Foto de dues espècies d’arcosaures actuals; un cocodril del Nil (Crocodylus niloticus) i un tàntal africà (Mycteria ibis). Foto de  Esquelet de la tortuga extingida Meiolania platyceps que visqué a Nova Caledònia fins fa 3000 anys. En aquesta foto s’aprecia el crani compacte i sense obertures temporals. Foto de
Esquelet de la tortuga extingida Meiolania platyceps que visqué a Nova Caledònia fins fa 3000 anys. En aquesta foto s’aprecia el crani compacte i sense obertures temporals. Foto de  Exemplar de tortuga lleopard (Stigmochelys pardalis) de Tanzània. Foto de
Exemplar de tortuga lleopard (Stigmochelys pardalis) de Tanzània. Foto de  Esquema
Esquema 



 En la fecundació, al ajuntar-se l’òvul i l’espermatozou (ambdós haploides, amb n nombre de cromosomes cadascun) s’obtindrà un individu amb material genètic únic, fruit de la combinació de l’ADN del pare i de la mare (diploide, amb dues còpies de cada cromosoma, 2n cromosomes a cada cèl·lula). Els animals triploides (3n) o tetraploides (4n) es donen en espècies híbrides asexuals, tot i que en molts casos no solen ser viables. Imatge de
En la fecundació, al ajuntar-se l’òvul i l’espermatozou (ambdós haploides, amb n nombre de cromosomes cadascun) s’obtindrà un individu amb material genètic únic, fruit de la combinació de l’ADN del pare i de la mare (diploide, amb dues còpies de cada cromosoma, 2n cromosomes a cada cèl·lula). Els animals triploides (3n) o tetraploides (4n) es donen en espècies híbrides asexuals, tot i que en molts casos no solen ser viables. Imatge de  Colònia d’abelles carnolianes (Apis mellifera carnica), una subespècie d’abella de la mel de l’est d’Europa. Foto de
Colònia d’abelles carnolianes (Apis mellifera carnica), una subespècie d’abella de la mel de l’est d’Europa. Foto de  Esquema del cicle vital d’un rotífer, en el que s’alterna la reproducció asexual amb partenogènesi en períodes de bones condicions ambientals, amb la reproducció sexual amb un mascle haploide quan les condicions no són òptimes. Imatge extreta de
Esquema del cicle vital d’un rotífer, en el que s’alterna la reproducció asexual amb partenogènesi en períodes de bones condicions ambientals, amb la reproducció sexual amb un mascle haploide quan les condicions no són òptimes. Imatge extreta de  Llangardaix cua de fuet de l’espècie Cnemidophorus uniparens que, com el seu nom indica, és una espècie partenogènica en que tots els exemplars són femelles. Foto de
Llangardaix cua de fuet de l’espècie Cnemidophorus uniparens que, com el seu nom indica, és una espècie partenogènica en que tots els exemplars són femelles. Foto de  Cria de dragó de Komodo (Varanus komodoensis) nascuda per partenogènesi accidental al Zoo de Chester. Foto de
Cria de dragó de Komodo (Varanus komodoensis) nascuda per partenogènesi accidental al Zoo de Chester. Foto de  Esquema de la formació dels cossos polars durant la oogènesi, els quals poden ajudar als rèptils partenogènics a recuperar la diploïdia. Esquema de
Esquema de la formació dels cossos polars durant la oogènesi, els quals poden ajudar als rèptils partenogènics a recuperar la diploïdia. Esquema de  Exemplar de serp cega de brahmin (Ramphotyphlops braminus), l’únic ofidi unisexual conegut, en el que tots els individus trobats fins ara són femelles. Foto extreta de
Exemplar de serp cega de brahmin (Ramphotyphlops braminus), l’únic ofidi unisexual conegut, en el que tots els individus trobats fins ara són femelles. Foto extreta de  Tres espècies de llangardaixos cua de fuet. El del mig, Cnemidophorus neomexicanus és una espècie unisexual partenogènica, resultat de l’hibridació de dues espècies bisexuals, C. inornatus (esquerra) i C. tigris (dreta). Foto de
Tres espècies de llangardaixos cua de fuet. El del mig, Cnemidophorus neomexicanus és una espècie unisexual partenogènica, resultat de l’hibridació de dues espècies bisexuals, C. inornatus (esquerra) i C. tigris (dreta). Foto de 

{kind=link}