

En Nueva Zelanda existe un reptil cuyo linaje surgió en la época de los dinosaurios. Aunque su aspecto externo se parece al de un lagarto, el tuatara (cuyo nombre significa “espalda espinosa” en lengua maorí) es un animal con muchas características únicas que hacen que se le clasifique en un orden propio separado del resto de reptiles. En esta entrada os explicaremos las principales características de esta reliquia del pasado tan interesante como amenazada.
ORIGEN Y EVOLUCIÓN
Los tuataras son unos reptiles inusuales cuyo linaje se remonta a hace 240 millones de años, a mitades del periodo Triásico. Los tuataras son lepidosaurios, aunque forman un linaje distinto a los escamosos, por lo que se encuentran en su propio orden, los rincocéfalos (orden Rhynchocephalia). Muchas especies florecieron durante el Mesozoico, aunque prácticamente todas fueron reemplazadas por los escamosos. A finales del Mesozoico solo quedaba una familia, los Sphenodontidae.

De todos los esfenodóntidos que existieron, solo los tuataras han sobrevivido hasta la actualidad. Tradicionalmente se consideraba que los tuataras incluían dos especies: el tuatara común (Sphenodon punctatus) y el tuatara de la Isla Brothers (Sphenodon guntheri), aunque análisis recientes han popularizado la idea de que el tuatara es una única especie, S. punctatus.
ANATOMÍA DEL TUATARA
Como ya hemos comentado, los tuataras se parecen externamente a un lagarto, teniendo cierto parecido con las iguanas. Los machos de tuatara son mayores que las hembras, llegando a los 61 cm de longitud y el quilo de peso, mientras que éstas solo alcanzan los 45 cm y el medio quilo. Los tuataras presentan una hilera de espinas en el dorso que les confiere su nombre común. Ésta es mayor en los machos, y puede erizarse para exhibirse.

Lo que realmente distingue a los tuataras es su anatomía interna. El resto de reptiles han modificado mucho la estructura de su cráneo, pero los tuataras han conservado la estructura diápsida original sin muchos cambios. Mientras que cocodrilos y tortugas han desarrollado cráneos macizos, los tuataras conservan amplias aperturas temporales, y aunque los escamosos han desarrollado cráneos y mandíbulas muy flexibles, los tuataras mantienen un cráneo rígido. Además, a diferencia de la mayoría de reptiles, los tuataras no presentan oídos externos.

El nombre Rhynchocephalia significa “cabeza de pico” y se refiere a la estructura de pico de su premaxilar. Los tuataras también son de los pocos reptiles con dientes acrodontos, los cuáles se encuentran fusionados al maxilar y la mandíbula y no se renuevan. Además, presentan un movimiento mandibular único tipo sierra, moviendo la mandíbula adelante y atrás.
Vídeo de YouOriginal, de unos tuataras en cautividad alimentándose. En este vídeo podemos apreciar el movimiento singular de la mandíbula.
Finalmente, una de las características anatómicas más increíbles de los tuataras es que éstos conservan el ojo parietal o pineal. Ésta es una estructura reminiscente de los primeros tetrápodos, conectada con la glándula pineal y que está involucrada en la regulación de la temperatura y los ritmos circadianos. Aunque algunos otros animales también lo conservan, los tuataras presentan un auténtico tercer ojo, con una retina y cristalino completos, aunque éste se va cubriendo de escamas con la edad.
HÁBITAT Y BIOLOGÍA
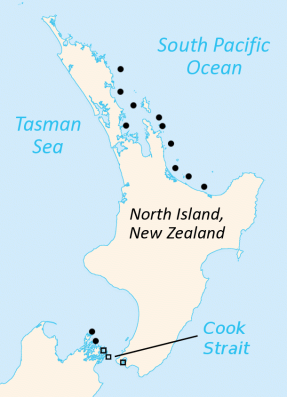
Los tuataras viven en unos treinta islotes en el estrecho de Cook, entre las dos islas principales de Nueva Zelanda. Además, la antigua especie S. guntheri se encuentra en la isla de Brothers, en la parte nororiental de isla Sur. Todas las poblaciones viven en zonas boscosas o de matorral costeras, con suelos blandos donde poder cavar. Además, en gran parte de su área de distribución existen colonias de aves marinas, cuyos nidos son aprovechados por los tuataras.

Comparados con la mayoría de reptiles, los tuataras viven en hábitats relativamente fríos, con temperaturas anuales que oscilan entre los 5 y los 28°C. Los tuataras son principalmente nocturnos, saliendo de sus madrigueras normalmente por la noche, aunque a veces se les puede encontrar tomando el sol a pleno día (especialmente en invierno).
Los tuataras tienen pocos depredadores naturales. Aparte de algunos animales introducidos, sólo las gaviotas y algunas aves de presa presentan un peligro para estos reptiles. Su dieta, en cambio, es bastante variada. Siendo depredadores que esperan a que sus presas les pasen por delante, los tuataras se alimentan principalmente de invertebrados como escarabajos, grillos y arañas, aunque puede llegar a depredar pequeños lagartos, huevos y polluelos de aves, e incluso a tuataras más pequeños. Como sus dientes acrodontos no se renuevan, éstos se van desgastando al cabo del tiempo, por lo que los ejemplares más viejos suelen alimentarse de presas más blandas como caracoles y gusanos.
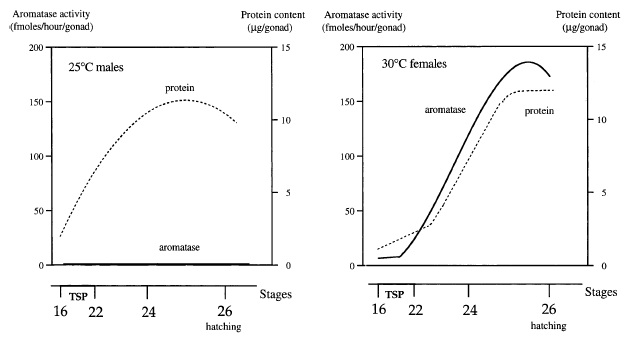
Los tuataras se reproducen entre enero y marzo (verano), momento en el que los territoriales machos compiten por las hembras, las cuáles pondrán unos 18-19 huevos entre octubre y diciembre (primavera). El sexo de las crías dependerá de la temperatura de incubación (machos a temperaturas más altas, hembras a más bajas). Los huevos eclosionarán al cabo de 11-16 meses (uno de los tiempos de incubación más largos de todos los reptiles), de los que saldrán pequeños tuataras que evitarán a los adultos caníbales siendo principalmente diurnos.
Vídeo único del nacimiento de un tuatara en la Victoria University de Wellington. La marca translúcida de la cabeza del pequeño tuatara corresponde al ojo parietal.
Como ya hemos visto por su largo período de incubación, los tuataras se desarrollan lentamente. Estos reptiles no llegarán a la madurez sexual hasta pasados los 12 años, aunque siguen creciendo a partir de entonces. Además, los tuataras son animales muy longevos, llegando a vivir hasta más de 60 años en estado salvaje. En cautividad pueden llegar a superar los 100 años de edad.
CONSERVACIÓN Y AMENAZAS
Antes de la llegada del hombre, los tuataras estaban presentes en las dos islas principales de Nueva Zelanda y en muchos más islotes. Cuando los colonos europeos llegaron, los tuataras ya sólo se encontraban en unas 32 pequeñas islas. Se cree que la desaparición de los tuataras de las islas principales se debe principalmente a la destrucción del hábitat y a la introducción de mamíferos foráneos como las ratas. Otras amenazas son la baja diversidad genética por el aislamiento de las distintas poblaciones y el cambio climático, que puede afectar al sexo de la descendencia.
Cuando el ser humano llegó a las islas, se cree que el 80% de Nueva Zelanda estaba cubierta de bosques. Con la llegada de las primeras tribus polinesias hacia el año 1250, empezó la deforestación de más de la mitad del archipiélago. Siglos después, con la llegada de los europeos, esta deforestación se intensificó aún más, hasta la situación actual, que solo se conserva el 23% del bosque original.

La introducción de mamíferos foráneos ha sido uno de los principales factores de declive de los tuataras en la actualidad, en especial la introducción de la rata del Pacífico (Rattus exulans). Este roedor ha afectado a las poblaciones, no solo de tuataras, sino también las de muchas especies de aves endémicas de Nueva Zelanda. En estudios de convivencia entre las ratas y los tuataras, se ha observado que las ratas, además de depredar sobre huevos y juveniles, también compiten con los tuataras adultos por los recursos. Con un ciclo vital tan lento, los tuataras no pueden recuperarse de este impacto.

Aun así, actualmente los tuataras están clasificados como bajo “preocupación menor” en la lista roja de la IUCN. Esto es gracias a los grandes esfuerzos de grupos conservacionistas que han contribuido a la recuperación de esta especie. Una de las principales tareas ha sido la eliminación de la rata del Pacífico de las principales islas donde habitan los tuataras. Para ello, se realizó un esfuerzo titánico en muchas islas en las que se capturaron poblaciones enteras de tuataras para la reproducción en cautividad, mientras se eliminaba a las ratas de dichas islas. Una vez eliminada su principal amenaza, todos los individuos capturados y sus descendientes nacidos en cautividad fueron devueltos a sus hábitats naturales para que pudieran vivir sin este fiero competidor.
Vídeo de Carla Braun-Elwert, sobre el éxito reproductor de una vieja pareja de tuataras.
Actualmente, la población salvaje de tuataras se estima entre los 60.000 y los 100.000 individuos. Se puede decir que este fósil viviente, que estuvo a punto de desaparecer después de millones de años de existencia, recibió una segunda oportunidad para seguir habitando las increíbles islas neozelandesas. Esperemos que en el futuro, podamos seguir disfrutando de la existencia de estos reptiles, únicos supervivientes de un linaje prácticamente extinto por muchos siglos más.
REFERENCIAS
Se han consultado las siguientes fuentes durante la elaboración de esta entrada:
- Halliday & Adler (2007). La gran enciclopedia de los Anfibios y Reptiles. Editorial Libsa.
- Arkive. Tuatara (Sphenodon punctatus).
- The Reptile Database. Sphenodon punctatus.
- Department of Conservation, Te Papa Atawhai. Tuatara.
- Imagen de portada de Michal Klajban.













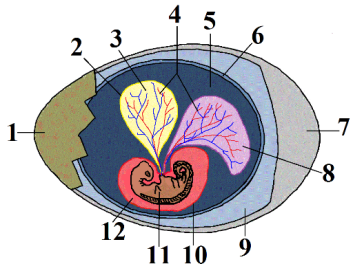 Esquema de l’ou d’un cocodril: 1. closca de l’ou 2. sac vitel·lí 3. vitel (nutrients) 4. vasos sanguinis 5. amni 6. cori 7. aire 8. alantoide 9. albúmina (clara de l’ou) 10. sac amniòtic 11. embrió 12. líquid amniòtic. Imatge de
Esquema de l’ou d’un cocodril: 1. closca de l’ou 2. sac vitel·lí 3. vitel (nutrients) 4. vasos sanguinis 5. amni 6. cori 7. aire 8. alantoide 9. albúmina (clara de l’ou) 10. sac amniòtic 11. embrió 12. líquid amniòtic. Imatge de 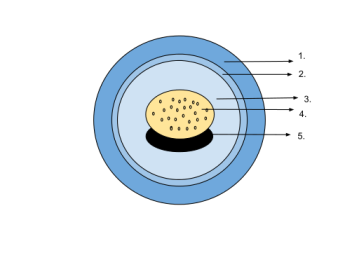 Esquema de l’ou d’un amfibi: 1. càpsula gelatinosa 2. membrana vitel·lina 3. fluid perivitel·lí 4. vitel 5. embrió. Imatge de
Esquema de l’ou d’un amfibi: 1. càpsula gelatinosa 2. membrana vitel·lina 3. fluid perivitel·lí 4. vitel 5. embrió. Imatge de  Reconstrucció de Solenodonsaurus janenschi, un dels candidats a ser el primer amniota, que visqué fa 320-305 milions d’anys a l’actual República Txeca. Recontrucció de
Reconstrucció de Solenodonsaurus janenschi, un dels candidats a ser el primer amniota, que visqué fa 320-305 milions d’anys a l’actual República Txeca. Recontrucció de 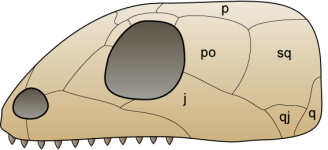 Esquema d’un crani anàpsid, de
Esquema d’un crani anàpsid, de 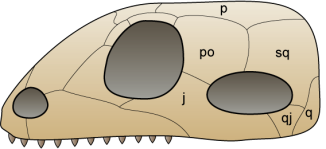 Esquema d’un crani sinàpsid, de
Esquema d’un crani sinàpsid, de 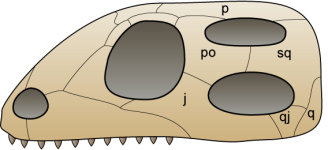 Esquema d’un crani diàpsid, de
Esquema d’un crani diàpsid, de 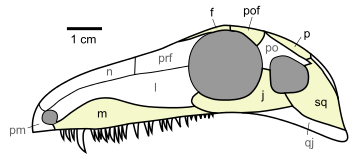 Dibuix del crani de Archaeothyris, el que es creu que va ser un dels primers sinàpsids que visqué fa uns 306 milions d’anys a Nova Escòcia. Dibuix de
Dibuix del crani de Archaeothyris, el que es creu que va ser un dels primers sinàpsids que visqué fa uns 306 milions d’anys a Nova Escòcia. Dibuix de  Reconstrucció de Dimetrodon grandis, un dels sinàpsids més coneguts, de fa uns 280 milions d’anys. Reconstrucció de
Reconstrucció de Dimetrodon grandis, un dels sinàpsids més coneguts, de fa uns 280 milions d’anys. Reconstrucció de 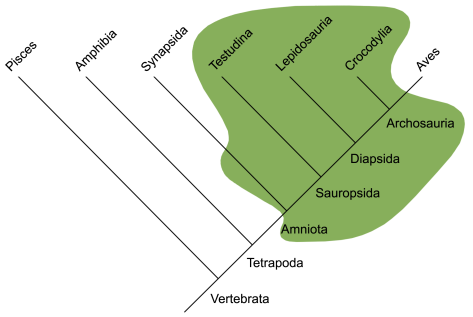 Arbre evolutiu dels vertebrats actuals, on es marca de color verd als grups antigament considerats rèptils. Com es veu, la concepció tradicional de “rèptil” inclou als avantpassats dels mamífers i exclou a les aus. Imatge de
Arbre evolutiu dels vertebrats actuals, on es marca de color verd als grups antigament considerats rèptils. Com es veu, la concepció tradicional de “rèptil” inclou als avantpassats dels mamífers i exclou a les aus. Imatge de  Muda sencera de la pell d’una serp rata. Foto de
Muda sencera de la pell d’una serp rata. Foto de  Foto d’una tuatara (Sphenodon punctatus), de
Foto d’una tuatara (Sphenodon punctatus), de  Fotos d’alguns escamosos d’esquerra a dreta i de dalt a baix: Iguana verda (Iguana iguana, de
Fotos d’alguns escamosos d’esquerra a dreta i de dalt a baix: Iguana verda (Iguana iguana, de 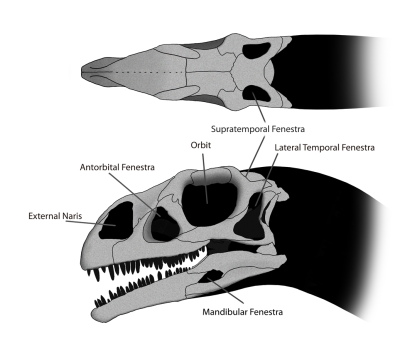 Dibuix del crani del dinosaure Massospondylus en el que es veuen les diferents obertures que caracteritzen als arcosaures diàpsids. Imatge de
Dibuix del crani del dinosaure Massospondylus en el que es veuen les diferents obertures que caracteritzen als arcosaures diàpsids. Imatge de  Foto de dues espècies d’arcosaures actuals; un cocodril del Nil (Crocodylus niloticus) i un tàntal africà (Mycteria ibis). Foto de
Foto de dues espècies d’arcosaures actuals; un cocodril del Nil (Crocodylus niloticus) i un tàntal africà (Mycteria ibis). Foto de  Esquelet de la tortuga extingida Meiolania platyceps que visqué a Nova Caledònia fins fa 3000 anys. En aquesta foto s’aprecia el crani compacte i sense obertures temporals. Foto de
Esquelet de la tortuga extingida Meiolania platyceps que visqué a Nova Caledònia fins fa 3000 anys. En aquesta foto s’aprecia el crani compacte i sense obertures temporals. Foto de  Exemplar de tortuga lleopard (Stigmochelys pardalis) de Tanzània. Foto de
Exemplar de tortuga lleopard (Stigmochelys pardalis) de Tanzània. Foto de 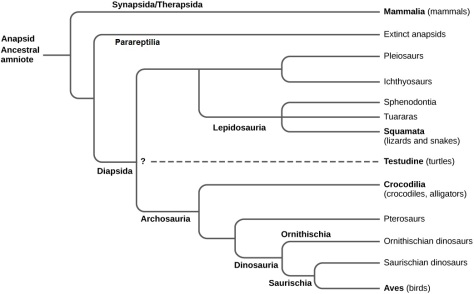 Esquema
Esquema 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}