

¿Cómo consiguen algunos insectos, arañas o lagartos caminar por paredes lisas e incluso boca abajo y no caerse? ¿Por qué, de ser real, Spiderman no podría engancharse en las paredes como lo hacen estos animales?
Científicos de diferentes áreas todavía buscan comprender los mecanismos que usan algunos animales para caminar sobre este tipo de superficies sin resbalarse o precipitarse. A continuación, te explicamos qué sabe la comunidad científica sobre este fenómeno.
Animales que caminan por la pared: un reto a la gravedad
La competencia por el espacio y los recursos (nicho ecológico) ha dado lugar a numerosas e increíbles adaptaciones a lo largo de la evolución, como la miniaturización.
Cuando una superficie es demasiado lisa, de manera que las uñas, las garras o las fuerzas de fricción resultan insuficientes para desplazarse sobre ella sin caerse, entran en juegos mecanismos de adhesión dinámica: aquellos que permiten al animal desplazarse sobre superficies verticales lisas o boca abajo enganchándose y desenganchándose rápidamente. La aparición de estructuras adhesivas dinámicas ha permitido a diversos animales explotar nuevos ambientes, pudiendo desplazarse para cazar o permanecer inmóviles el tiempo necesario para huir de sus depredadores allí donde la mayoría no podría estar estable más que unos pocos segundos.

El desarrollo de estructuras adhesivas dinámicas en las extremidades es típico de insectos y de arañas, de algunos reptiles como los geckos y ciertas lagartijas, y de anfibios como las ranas arborícolas. Puntualmente, también se ha observado en pequeños mamíferos como murciélagos y pósums, unos marsupiales arborícolas procedentes de Australia y de ciertas regiones del sudeste asiático.
El hecho de que grupos tan diferentes de animales presenten una adaptación similar se explica por un proceso de convergencia evolutiva: ante un mismo problema (competencia por el espacio y los recursos, elevada presión de depredación, etc.), la evolución tiende a soluciones iguales o similares (estructuras adhesivas para acceder a otros espacios).
Los límites de la adaptación (o por qué Spiderman no podría caminar por las paredes)
Estudiar el mecanismo mediante el cual algunos animales caminan sobre superficies verticales lisas o invertidas es clave para el desarrollo industrial de nuevas y más potentes sustancias adhesivas. No es de extrañar, por lo tanto, que haya muchos estudios al respecto.
¿Podrá el ser humano escalar paredes como lo hace Spiderman algún día? Labonte et al. (2016) nos explica por qué Spiderman como tal no podría existir. O, al menos, cómo debería ser realmente para poder adherirse a las paredes como una araña.

Sin entrar en las estrategias propias de cada organismo (de las cuales hablaremos después), el principio básico por el cual insectos, arañas o geckos pueden caminar sobre superficies verticales lisas o boca abajo es su relación superficie/volumen: a menor tamaño del animal, mayor es la superficie de su cuerpo respecto a su volumen y menor la cantidad de superficie adhesiva necesaria para poder desplazarse sin caerse debido al peso. Así pues, los geckos serían los animales conocidos con el tamaño más grande (relación superficie/volumen más pequeña) capaces de caminar sobre superficies verticales lisas o boca abajo sin sufrir modificaciones anatómicas que harían inviable su desarrollo.
¿Y qué significa “sin sufrir modificaciones anatómicas”? Los mismos autores explican que a mayor tamaño del animal, mayor es la superficie adhesiva necesaria para desplazarse sin desprenderse. El crecimiento de la superficie adhesiva con respecto al tamaño del animal sigue un patrón de alometría positiva extrema: por un pequeño incremento del tamaño del animal, se produce un aumento significativamente mayor de la superficie adhesiva. Según este estudio, la superficie adherente respecto a la superficie total puede ser hasta 200 veces mayor en geckos que en ácaros.

Sin embargo, la misma alometría se rige por una serie de constricciones (limitaciones) anatómicas. Así, para que existiera un animal de mayor tamaño que un gecko capaz de caminar sobre una superficie vertical lisa o invertida, éste debería desarrollar, por ejemplo, unas extremidades enormes con una superficie adherente igualmente grande. Si bien pudiera tener sentido desde un punto de vista físico, las constricciones anatómicas hacen inviable la existencia de animales con estas características.
Ahora ya podemos responder la pregunta “¿Por qué Spiderman no podría adherirse a las paredes?”. Según este estudio, para que un ser humano pudiera caminar por las paredes como una araña su cuerpo debería estar recubierto al menos de un 40% de estructuras adhesivas (un 80% si contamos únicamente su parte frontal); o eso, o tener brazos o piernas absurdamente grandes e imposibles desde un punto de vista anatómico.
Gran diversidad de estrategias
La adhesión dinámica debe ser suficientemente fuerte para que el animal no caiga al estar quieto, pero suficientemente débil para poder desengancharse sin problemas al dar un paso.
Para conseguirlo, existen diferentes estrategias.

1) Adhesión húmeda
Interviene una sustancia líquida.
Insectos
Los insectos presentan dos sistemas:
Patas con almohadillas lisas: lo encontramos, por ejemplo, en hormigas, abejas, cucarachas y saltamontes. El último segmento de sus patas (pretarso), las uñas o las tíbias presentan una o varias almohadillas extremadamente blandas y deformables (como los arolios en el pretarso). A pequeña escala, ninguna superficie es totalmente lisa, por lo que estas almohadillas se deforman hasta ocupar todos sus espacios disponibles.

Patas con almohadillas peludas: lo encontramos en escarabajos y moscas, entre otros. Las almohadillas de estos insectos están densamente cubiertas de pequeñas estructuras similares a pelos, las setas, gracias a las cuales el contacto con la superficie aumenta.

En ambos casos, interviene un líquido con una fase hidrofóbica y otra hidrofílica. Estudios con hormigas han demostrado que las terminaciones de sus patas secretan una fina capa de líquido que incrementa el contacto entre el pretarso y la superficie sobre la que caminan, rellenando los huecos restantes y actuando como un adhesivo bajo los principios de capilaridad (tensión superficial) y viscosidad.
Si queréis conocer más a fondo este mecanismo, ¡no os perdáis este increíble vídeo sobre las hormigas!:
Ranas arborícolas
Las almohadillas de los dedos de las ranas arborícolas están compuestas de células epiteliales columnares separadas entre sí. Entre ellas, numerosas glándulas vierten una sustancia mucosa a los espacios existentes. La separación de las células permite, por una parte, que las almohadillas se deformen para adaptarse al terreno y, por otra, que la mucosidad circule entre ellas y asegure la adhesión. Además, en ambientes húmedos (muchas de estas ranas viven en selvas), estos espacios facilitan la eliminación del exceso de agua que las haría resbalar.

En el siguiente vídeo, puedes apreciar con más detalle las patas de una de las ranas arborícolas más conocidas:
Las ranas arborícolas presentan un sistema similar al de almohadillas lisas de los insectos. De hecho, a muchos aumentos las microestructuras adhesivas en grillos y ranas es prácticamente idéntica. Esto llevó a Barnes (2007) a considerar la adhesión húmeda como una de las más exitosas.

Pósums
Los estudios más detallados se han realizado sobre el pósum pigmeo acróbata (Acrobates pygmaeus), un pequeño marsupial del tamaño de un ratón capaz de escalar superficies de vidrio usando las grandes almohadillas de sus patas. Estas almohadillas están compuestas de múltiples capas de células epiteliales escamosas separadas por surcos que facilitan su deformación y por los que circula el sudor, que es el líquido que usan para adherirse.


2) Adhesión seca
No intervienen líquidos.
Arañas y geckos
Tanto arañas como geckos se rigen por el mismo principio de adhesión: las fuerzas de Van de Waals. A diferencia de insectos, ranas y pósums, no segregan líquidos adhesivos.
Las fuerzas de Van der Waals resultan de la interacción entre moléculas o átomos sin que exista un enlace químico entre ellos, y su energía depende de la distancia. Estas interacciones aparecen entre los “pelos” o setas de las palmas de los geckos (las cuales están surcadas por pliegues, las lamelas) y las setas de las patas de las arañas (que están cubiertas de muchas pilosidades formando las escópulas), y la superficie sobre la que caminan.


Estudios recientes, sin embargo, sugieren que la adhesión en los geckos no se debería principalmente a estas fuerzas, sino a las interacciones electrostáticas (diferente polaridad entre las setas y la superficie), tras comprobar que su capacidad adhesiva menguaba sobre materiales menos energéticos, como el teflón.
Sea como sea, la habilidad de los geckos para trepar es impresionante. Sino, mira este vídeo del gran David Attenborough:
Succión
Murciélagos
Los murciélagos de ventosas (familia Thyropteridae), originarios de Centroamérica y el norte de Sudamérica, presentan unas ventosas en forma de disco en sus pulgares y en la planta de las patas traseras que les permiten desplazarse sobre superficies lisas. En el interior de estos discos, la presión se reduce y el murciélago queda adherido por succión. De hecho, un solo disco puede soportar el peso de todo el animal.

Después de conocer todas estas estrategias, ¿creéis que Spiderman está a la altura?
Imagen de portada de autor desconocido. Fuente: link.

























 El veneno de muchos animales es útil tanto para el desarrollo de antídotos, como para la investigación de analgésicos y otros medicamentos. Foto de la extracción de veneno de una víbora gariba (Echis carinatus), de
El veneno de muchos animales es útil tanto para el desarrollo de antídotos, como para la investigación de analgésicos y otros medicamentos. Foto de la extracción de veneno de una víbora gariba (Echis carinatus), de  Serpiente lobo de la India (Lycodon aulicus), ejemplo de ofidio. Foto de
Serpiente lobo de la India (Lycodon aulicus), ejemplo de ofidio. Foto de  Basilisco marrón (Basiliscus vittatus), ejemplo de iguanio. Foto de
Basilisco marrón (Basiliscus vittatus), ejemplo de iguanio. Foto de  Varano sordo de Borneo (Lanthanotus borneensis), ejemplo de anguimorfo. Foto de
Varano sordo de Borneo (Lanthanotus borneensis), ejemplo de anguimorfo. Foto de  Cráneo de helodermátido, en el que observamos los afilados dientes con los que inyectan el veneno. Imagen de
Cráneo de helodermátido, en el que observamos los afilados dientes con los que inyectan el veneno. Imagen de  Foto de
Foto de  Foto de un dragón barbudo del este, en la que vemos el interior de su boca de color amarillo. ¿Nos estará intentando avisar de algo con esta coloración? Foto de
Foto de un dragón barbudo del este, en la que vemos el interior de su boca de color amarillo. ¿Nos estará intentando avisar de algo con esta coloración? Foto de  Sección transversal de la boca de un dragón barbudo del este, donde se ven las glándulas venenosas incipientes tanto de la mandíbula superior (mxivg) como de la inferior (mnivg). Imagen extraída de
Sección transversal de la boca de un dragón barbudo del este, donde se ven las glándulas venenosas incipientes tanto de la mandíbula superior (mxivg) como de la inferior (mnivg). Imagen extraída de  Varano gigante australiano o “perentie” (Varanus giganteus) un varánido típico, con cuello largo, patas robusta, metabolismo activo y sentidos desarrollados. Foto de
Varano gigante australiano o “perentie” (Varanus giganteus) un varánido típico, con cuello largo, patas robusta, metabolismo activo y sentidos desarrollados. Foto de  Foto donde vemos la temida saliva de los varanos, concretamente de un varano acuático (Varanus salvator). Imagen de
Foto donde vemos la temida saliva de los varanos, concretamente de un varano acuático (Varanus salvator). Imagen de  Cráneo de megalania (Varanus priscus) en el que vemos los dientes sin surcos. Este varano extinto de más de 5 metros de largo, fue el animal venenoso más grande conocido.
Cráneo de megalania (Varanus priscus) en el que vemos los dientes sin surcos. Este varano extinto de más de 5 metros de largo, fue el animal venenoso más grande conocido.  Grupo de varanos de Komodo (Varanus komodoensis) devorando un cerdo recién cazado. Imagen extraída de
Grupo de varanos de Komodo (Varanus komodoensis) devorando un cerdo recién cazado. Imagen extraída de 

 Imagen de microscopio de un corte transversal del cráneo de una serpiente, en la que se ve el epitelio olfativo, tanto de la cavidad nasal como del órgano vomeronasal. Imagen de Elliott Jacobson.
Imagen de microscopio de un corte transversal del cráneo de una serpiente, en la que se ve el epitelio olfativo, tanto de la cavidad nasal como del órgano vomeronasal. Imagen de Elliott Jacobson. Esquema de la posición del órgano vomeronasal. Éste se forma durante el desarrollo embrionario a partir de la cavidad nasal y tiene una apertura al paladar. Imagen de
Esquema de la posición del órgano vomeronasal. Éste se forma durante el desarrollo embrionario a partir de la cavidad nasal y tiene una apertura al paladar. Imagen de  Los varanos (parientes de las serpientes) también presentan una lengua bífida que les permite oler el aire. Foto de un varano acuático (Varanus salvator) en la India, de
Los varanos (parientes de las serpientes) también presentan una lengua bífida que les permite oler el aire. Foto de un varano acuático (Varanus salvator) en la India, de  Foto de una
Foto de una  Retrato de una pitón de la India (Python molurus) en el que se ve claramente la ausencia de oído externo. Foto de
Retrato de una pitón de la India (Python molurus) en el que se ve claramente la ausencia de oído externo. Foto de 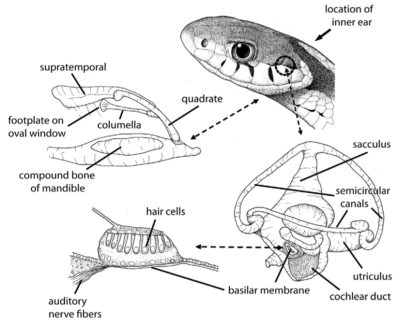 Esquema del aparato auditivo de una serpiente cualquiera. Imagen de Dan Dourson.
Esquema del aparato auditivo de una serpiente cualquiera. Imagen de Dan Dourson. Las serpientes acuáticas como la anaconda (Eunectes murinus) pueden detectar con todo el cuerpo los sonidos de animales moviéndose bajo el agua. Foto de
Las serpientes acuáticas como la anaconda (Eunectes murinus) pueden detectar con todo el cuerpo los sonidos de animales moviéndose bajo el agua. Foto de  La víbora cornuda arábica (Cerastes gasperettii) es una serpiente que
La víbora cornuda arábica (Cerastes gasperettii) es una serpiente que  Los ofidios más primitivos, como esta serpiente ciega europea (Typhlops vermicularis), tienen ojos pequeños y poco desarrollados. Imagen de
Los ofidios más primitivos, como esta serpiente ciega europea (Typhlops vermicularis), tienen ojos pequeños y poco desarrollados. Imagen de  Víbora ratonera (Pantherophis obsoletus) a punto de mudar de piel, momento en que la escama ocular se ve opaca. Foto de
Víbora ratonera (Pantherophis obsoletus) a punto de mudar de piel, momento en que la escama ocular se ve opaca. Foto de  Muchas serpientes arborícolas como esta serpiente látigo verde (Ahaetulla nasuta) presentan pupilas horizontales que les permiten aumentar su campo de visión, haciendo que puedan calcular mejor las distancias entre rama y rama. Foto de
Muchas serpientes arborícolas como esta serpiente látigo verde (Ahaetulla nasuta) presentan pupilas horizontales que les permiten aumentar su campo de visión, haciendo que puedan calcular mejor las distancias entre rama y rama. Foto de 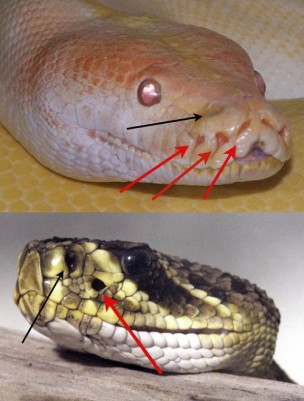 Fotos de una pitón y un crótalo donde se señalan tanto las narinas (flechas negras) como las cavidades termorreceptoras (flechas rojas). Imagen de
Fotos de una pitón y un crótalo donde se señalan tanto las narinas (flechas negras) como las cavidades termorreceptoras (flechas rojas). Imagen de  Esquema de la estructura de la cavidad termorreceptora de un crótalo. Ésta presenta una membrana sensible a los cambios de temperatura, detrás de la cual hay una cámara con aire y nervios sensibles al calor. Este aire se dilata con los incrementos de temperatura y activa el nervio trigémino. Imagen de
Esquema de la estructura de la cavidad termorreceptora de un crótalo. Ésta presenta una membrana sensible a los cambios de temperatura, detrás de la cual hay una cámara con aire y nervios sensibles al calor. Este aire se dilata con los incrementos de temperatura y activa el nervio trigémino. Imagen de 

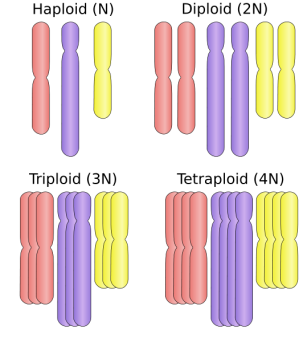 En la fecundación, al unirse el óvulo y el espermatozoide (ambos haploides, con n número de cromosomas cada uno) se obtendrá un individuo con material genético único, fruto de la combinación del ADN del padre y de la madre (diploide, con dos copias de cada cromosoma, 2n cromosomas en cada célula). Los animales triploides (3n) o tetraploides (4n) se dan es especies híbridas asexuales, aunque en muchos casos no suelen ser viables. Imagen de
En la fecundación, al unirse el óvulo y el espermatozoide (ambos haploides, con n número de cromosomas cada uno) se obtendrá un individuo con material genético único, fruto de la combinación del ADN del padre y de la madre (diploide, con dos copias de cada cromosoma, 2n cromosomas en cada célula). Los animales triploides (3n) o tetraploides (4n) se dan es especies híbridas asexuales, aunque en muchos casos no suelen ser viables. Imagen de  Colonia de abejas carnolianas (Apis mellifera carnica), una subespecie de abeja de la miel del este de Europla. Foto de
Colonia de abejas carnolianas (Apis mellifera carnica), una subespecie de abeja de la miel del este de Europla. Foto de  Esquema del ciclo vital de un rotífero, en el que se alterna la reproducción asexual con partenogénesis en períodos de buenas condiciones ambientales, con la reproducción sexual con un macho haploide cuando las condiciones no son óptimas. Imagen extraída de
Esquema del ciclo vital de un rotífero, en el que se alterna la reproducción asexual con partenogénesis en períodos de buenas condiciones ambientales, con la reproducción sexual con un macho haploide cuando las condiciones no son óptimas. Imagen extraída de  Lagarto de cola látigo de la especie Cnemidophorus uniparens que, como su nombre indica, es una especie partenogénica en que todos los ejemplares son hembras. Foto de
Lagarto de cola látigo de la especie Cnemidophorus uniparens que, como su nombre indica, es una especie partenogénica en que todos los ejemplares son hembras. Foto de  Cría de dragón de Komodo (Varanus komodoensis) nacida por partenogénesis accidental en el Zoo de Chester. Foto de
Cría de dragón de Komodo (Varanus komodoensis) nacida por partenogénesis accidental en el Zoo de Chester. Foto de  Esquema de la formación de los cuerpos polares durante la ovogénesis, los cuales pueden ayudar a los reptiles partenogénicos a recuperar la diploidía. Esquema de
Esquema de la formación de los cuerpos polares durante la ovogénesis, los cuales pueden ayudar a los reptiles partenogénicos a recuperar la diploidía. Esquema de  Ejemplar de serpiente ciega de brahmin (Ramphotyphlops braminus), el único ofidio unisexual conocido, en el que todos los individuos encontrados hasta la fecha son hembras. Foto extraída de
Ejemplar de serpiente ciega de brahmin (Ramphotyphlops braminus), el único ofidio unisexual conocido, en el que todos los individuos encontrados hasta la fecha son hembras. Foto extraída de  Tres especies de lagartos de cola látigo. El de enmedio, Cnemidophorus neomexicanus es una especie unisexual partenogénica, resultado de la hibridación de dos especies bisexuales, C. inornatus (izquierda) y C. tigris (derecha). Foto de
Tres especies de lagartos de cola látigo. El de enmedio, Cnemidophorus neomexicanus es una especie unisexual partenogénica, resultado de la hibridación de dos especies bisexuales, C. inornatus (izquierda) y C. tigris (derecha). Foto de 
 Imagen de
Imagen de  Foto de un ratón espinoso del Cairo (Acomys cahirinus), un mamífero que es capaz de desprenderse de su cola y regenerarla. Foto de
Foto de un ratón espinoso del Cairo (Acomys cahirinus), un mamífero que es capaz de desprenderse de su cola y regenerarla. Foto de 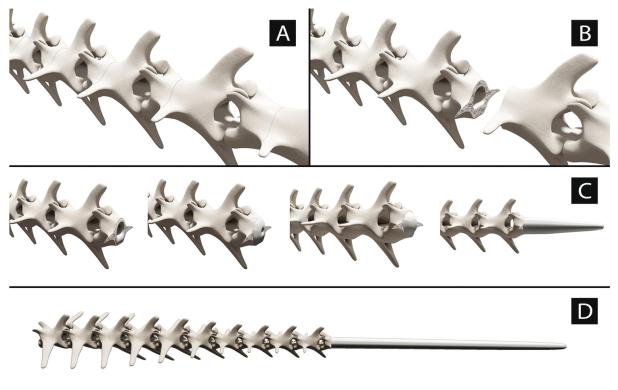 Modelo tridimensional de los planos de fractura de la cola de un lagarto y la regeneración post-autotomía de un tubo cartilaginoso. Imagen extraída de
Modelo tridimensional de los planos de fractura de la cola de un lagarto y la regeneración post-autotomía de un tubo cartilaginoso. Imagen extraída de  Foto de una lagartija parda (Podarcis liolepis) que ha perdido la cola. Foto de
Foto de una lagartija parda (Podarcis liolepis) que ha perdido la cola. Foto de  Detalle de la cola de una salamanquesa común (Tarentola mauritanica) que ha regenerado la cola sin acabar de perder la cola original. Foto de
Detalle de la cola de una salamanquesa común (Tarentola mauritanica) que ha regenerado la cola sin acabar de perder la cola original. Foto de  Renacuajo de rana de bosque (Rana sylvatica) que, cómo en todos los anfibios, pospone el desarrollo de las extremidades hasta el momento de la metamorfosis. Foto de
Renacuajo de rana de bosque (Rana sylvatica) que, cómo en todos los anfibios, pospone el desarrollo de las extremidades hasta el momento de la metamorfosis. Foto de  Esquema de la formación del blastema en el pez zebra (Danio rerio) otro organismo modelo. Imagen de
Esquema de la formación del blastema en el pez zebra (Danio rerio) otro organismo modelo. Imagen de  Foto de un ajolote, por
Foto de un ajolote, por 





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}